ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ОРГАНИЧЕСКОГО МИРА
1. Введение
2. Самопроизвольное зарождение
3. Как же возникла жизнь?
4. Развитие жизни на Земле (краткий очерк)
5. Эволюционная теория: драма в биологии
6. Приговор… из Библии
7. Список литературы
ВВЕДЕНИЕ
На протяжении тысячелетий людям казалось очевидным, что живая природа была создана такой, какой мы ее знаем сейчас, и всегда оставалась неизменной.
Но уже в глубокой древности высказывались догадки о постепенном изменении, развитии (эволюции) живой природы. Одним из предтеч эволюционных идей можно назвать древнегреческого философа Гераклита (VI - V вв. до н. э.), который сформулировал положение о постоянно происходящих в природе изменениях («все течет, все изменяется»).
Другой древнегреческий мыслитель - Эмпедокл - в V в. до н. э. выдвинул, вероятно, одну из древнейших теорий эволюции. Он считал, что вначале на свет появились разрозненные части различных организмов (головы, туловища, ноги). Они соединились между собой в самых невероятных сочетаниях. Так появились, в частности, кентавры (мифические полулюди - полукони). Позднее будто бы все нежизнеспособные комбинации погибли.
Великий древнегреческий ученый Аристотель выстроил все известные ему организмы в ряд по мере их усложнения. В XVII в. эту идею развил швейцарский натуралист Шарль Бонне, создав учение о «лестнице природы». На первой ступени «лестницы» находились «тонкие материи» - огонь, воздух, вода, земля; на следующих - растения и животные по степени сложности их строения, на одной из верхних ступеней - человек, а еще выше - небесное воинство и Бог. Правда, о возможности перехода «со ступени на ступень» речи, конечно, не шло, и к эволюции эта система имеет еще весьма отдаленное отношение.
Первую последовательную теорию эволюции живых организмов разработал французский ученый Жан Батист Ламарк в книге «Философия зоологии», вышедшей в 1809 г. Ламарк предположил, что в течение жизни каждая особь изменяется, приспосабливается к окружающей среде. Приобретенные ею на протяжении всей жизни новые признаки передаются потомству. Так из поколения в поколение накапливаются изменения. Но рассуждения Ламарка содержали ошибку, которая заключалась в простом факте: приобретенные признаки не наследуются. В конце XIX в. немецкий биолог Август Вейсман поставил известный эксперимент - на протяжении 22 поколений отрезал хвосты подопытным мышам. И все равно новорожденные мышата имели хвосты ничуть не короче, чем их предки.
Английский ученый Чарлз Дарвин в отличие от Ламарка обратил внимание на то, что хотя любое живое существо изменяется в течение жизни, но и рождаются особи одного вида неодинаковыми. Дарвин писал, что опытный фермер различает каждую из овец даже в большом по численности стаде. Например, шерсть их может быть светлее или темнее, гуще или реже и т. п. В обычных условиях среды такие различия несущественны. Но при перемене условий жизни эти мелкие наследственные изменения могут давать преимущества их обладателям. Среди множества бесполезных и вредных изменений могут встречаться и полезные.
Рассуждая таким образом, Дарвин пришел к идее естественного отбора. Особи с полезными отличиями лучше выживают и размножаются, передают свои признаки потомству. Поэтому в следующем поколении процент таких особей станет больше, через поколение - еще больше и т. д. Таков механизм эволюции. Дарвин писал: «Можно сказать, что естественный отбор ежедневно и ежечасно расследует по всему свету мельчайшие изменения, отбрасывая дурные, сохраняя и слагая хорошие, работая неслышно и невидимо…»
Эволюция разных видов идет с разной скоростью. К примеру, беспозвоночные, относящиеся к типу плеченогих, почти не изменились за последние 440 млн. лет. А в роде Человек, по данным палеонтологов, за последние 2 млн. лет возникло и вымерло несколько видов.
Конечно, взгляды на теорию эволюции не остались неизменными со времен Дарвина. К примеру, Дарвин счел очень серьезным возражение против своей теории, выдвинутое английским инженером Ф. Дженкином (оно получило название «кошмара Дженкина»). Дженкин рассуждал так: допустим, у одной особи случайно появился какой-то полезный признак. Но у ее потомства этот признак «разбавится» ровно вдвое, у следующего поколения - еще более уменьшится, пока совершенно не «растворится» и не будет утрачен. В то время считалось (так думал и Дарвин), что у потомства признаки родителей могут сливаться (скажем, у белых и черных мышей родится потомство серого цвета). Это распространенное заблуждение опровергли только открытия Грегора Менделя, которые Дарвину еще не были известны.
В своей аргументации Дарвин опирался на множество примеров искусственного, проводимого человеком отбора (с помощью которого были созданы многие породы домашних животных и культурных растений). Но Дарвин не сумел представить ни одного убедительного примера происходящего в природе естественного отбора. Такие примеры были описаны учеными только в XX в. Самый известный из этих примеров - с бабочкой березовой пяденицей в Англии. Осматривая в 1950 г. коллекции бабочек, собранные за предшествующие сто лет, биологи обнаружили, что бабочки с черными крыльями встречались все чаще, а с серыми - все реже. Оказывается, днем пяденицы неподвижно сидят на стволах деревьев, полагаясь на свою маскирующую окраску. В XIX в. серая окраска превосходно скрывала бабочек на фоне лишайников, которыми были покрыты деревья. Но по мере того как загрязнение воздуха в Англии усиливалось, лишайники вымирали, а стволы становились черными от копоти. На темном фоне серые бабочки стали заметными для своих главных врагов - птиц. Черная же форма оказалась хорошо замаскированной. В результате соотношение черных и серых бабочек неуклонно изменялось в пользу черных. (Отметим, что единицей эволюции всегда является не особь, а популяция, т. е. Группа особей (в данном случае - пядениц), обитающих рядом друг с другом и скрещивающихся между собой).
Еще более яркий пример естественного отбора - возникновение устойчивости к ядохимикатам у насекомых. Профессор Кэролл Уильямс писал, что в начале 40-х гг. XX в. «в руках человека оказалось мощное оружие. Это был ядохимикат ДДТ, который, как всемогущий ангел-мститель, обрушивался на вредных насекомых. После первого же соприкосновения с ним комары, мухи, почти все насекомые срывались в штопор, падали, час-другой жужжали, лежа на спине, а потом погибали». Первые сообщения об устойчивости насекомых к ДДТ появились в 1947 г. и касались комнатной мухи. Из полчищ вредных насекомых систематически выживали лишь немногие, случайно оказавшихся более устойчивыми к яду. Но каждый следующий год в живых оставалось все более и более стойкое потомство. «Несколько лет спустя, - писал Уильямс, - комары, блохи, мухи и другие насекомые уже перестали обращать внимание на ДДТ. Скоро они начали его усваивать, потом полюбили». Такая устойчивость была обнаружена более чем у 200 видов насекомых, и список этот продолжал расти.
Совершенно аналогична история «привыкания» болезнетворных бактерий к антибиотикам и многим другим лекарствам.
САМОПРОИЗВОЛЬНОЕ ЗАРОЖДЕНИЕ
В течение долгих веков, свято веря в акт Божественного творения, люди, кроме того, были твердо убеждены, что жизнь постоянно зарождается самопроизвольно.
Еще древнегреческий философ Аристотель писал, что не только растения, черви, насекомые, но даже рыбы, лягушки и мыши могут рождаться из влажной почвы или гниющего ила.
Голландский ученый Ян ван Гельмонт в XVII в. описал свой опыт, утверждая, что живые мыши якобы зарождались у него из грязного белья и горсти пшеницы, запертых в шкафу.
Другой натуралист, Гриндель фон Ах, так рассказывал о якобы наблюдавшемся им самозарождении живой лягушки: «Хочу описать появление на свет лягушки, которое мне удалось наблюдать при помощи микроскопа. Однажды я взял каплю майской росы, и тщательно наблюдая за ней под микроскопом, заметил, что у меня сформировывается какое-то существо. Прилежно наблюдая на второй день, я заметил, что появилось уже туловище, но голова еще казалась не ясно сформированной; продолжая свои наблюдения на третий день, я убедился, что наблюдаемое мною существо есть ни что иное, как лягушка с головой и ногами. Прилагаемый рисунок все объясняет».
В 1688 г. итальянский ученый Франческо Реди решил проверить идею самопроизвольного зарождения жизни. Он рассказывал о своем опыте: «Я взял четыре сосуда, поместил в один из них мертвую змею, в другой - немного рыбы, в третий - дохлых угрей, в четвертый - кусок телятины, плотно закрыл их и запечатал. Затем я поместил то же самое в четыре других сосуда, оставив их открытыми. Вскоре мясо и рыба в открытых сосудах зачервивели. Можно было видеть, как мухи свободно залетают в сосуды и вылетают из них. Но в запечатанных сосудах я не увидел ни одного червяка, хотя прошло много дней после того, как в них была положена дохлая рыба и мясо».
В 1675 г. итальянский ученый Ладзаро Спаланцани прокипятил в запаянном сосуде крепкий мясной бульон. Прошло несколько дней, но никаких признаков жизни в бульоне не обнаружилось.
Наконец, в 1860 г. Луи Пастер с помощью ряда блестящих опытов, похожих на опыт Спалланцани, окончательно доказал, что жизнь в современных условиях не самозарождается. Он показал, что даже бактерии могут возникать только от других бактерий.
КАК ЖЕ ВОЗНИКЛА ЖИЗНЬ?
Опыты Пастера не разрешили вопрос о происхождении жизни, а поставили его с новой остротой. Если жизнь в современных условиях не самозарождается, то когда и как она возникла впервые?
Наблюдаемая нами Вселенная, по данным современной науки, возникла в результате Большого Взрыва около 15-20 млрд. лет назад. Возраст нашей планеты - около 5 млрд. лет. Сейчас большинство ученых склоняется к мнению о том, что жизнь зародилась на Земле на заре ее существования.
Древнейшая Земля весьма мало напоминала планету, на которой мы живем. Ее атмосфера состояла из водяных паров, углекислого газа и, по одним данным, - из азота, по другим - из метана и аммиака. Кислорода в воздухе безжизненной планеты не было. И, надо сказать, отсутствие кислорода было необходимо для возникновения жизни. Возможно, что необычное словосочетание «смертоносный кислород» вызовет некоторое удивление. Между тем кислород разрушительно действует на органические молекулы. Мы привыкли к его воздействию, но на Земле и сейчас есть бактерии, которые воспринимают кислород как яд и в его присутствии жить не могут. Кислородная атмосфера делает невозможным в наше время самозарождение жизни.
Итак, в атмосфере древней Земли гремели грозы, ее пронизывало жестокое ультрафиолетовое излучение Солнца, на планете извергались вулканы.
Под влиянием всех этих воздействий в первичном океане, покрывавшем поверхность Земли, образовывались органические вещества - простейшие «кирпичики», из которых строится все живое. В наше время их немедленно поглотили бы бактерии и грибы. Но тогда их еще не было, и поэтому органические вещества накапливались, пока весь первичный океан не превратился в «теплый разбавленный бульон».
Такое предположение впервые высказал в 1922 г. советский биолог Александр Опарин.
В 1953 г. американский биолог Стэнли Миллер решил проверить гипотезу Опарина и воспроизвел в специальной установке природные условия древней Земли. В стеклянном сосуде находились нагретая вода («океан») и смесь газов - аммиака, метана и водорода («первичная атмосфера»). Через «атмосферу» проскакивали искры - «молнии». Опыт продолжался в течении недели.
Через неделю «первичный бульон» проанализировали и нашли в нем многие органические вещества, в том числе 5 аминокислот. В другой раз в результате такого же опыта были обнаружены даже нуклеиновые кислоты - цепочки, длиной до шести звеньев.
Согласно одной гипотезе, содержание органических веществ выше всего было в высыхающих лужах, оставшихся на берегу океана после отлива. Здесь образовывались цепочки белков и нуклеиновых кислот. При этом чем длиннее была цепочка, тем она была устойчивее. Она закручивалась в клубок, который разрушался уже не так легко.
Опарин считал, что главная роль в превращении неживого в живое принадлежала белкам. В «первичном бульоне» образовывались «сгустки» белка «коацерваты). Они могли вбирать в себя новые питательные вещества, разбиваться на более мелкие капельки. Конечно, они еще не были живыми.
По словам Опарина, расстояние от этих «сгустков» до самых примитивных бактерий ничуть не меньше, чем от амебы до человека. Главное, что отличало «сгустки» от клеток, - неспособность точно воспроизводить самих себя.
Чтобы «штамповать» одинаковые белки, нужна матрица. В ныне живущих организмах (от бактерий и вирусов до человека) этой матрицей служат нуклеиновые кислоты (РНК, ДНК).
В какой момент белковые «сгустки» «перешагнули» порог живого? Тогда, когда включили в себя нуклеиновые кислоты, которые позволили создавать хотя бы грубые, приблизительные копии уже имеющихся белков. Это были уже зачатки примитивных клеток.
Один из скептиков высказал мнение, что возникновение жизни в результате перечисленных процессов столь же неправдоподобно, как сборка самолета «Боинг-747» в результате урагана, пронесшегося над мусорной свалкой. Но не будем забывать, что на протяжении длительного времени (миллиарда лет) в огромном пространстве, где происходил «опыт» (весь земной океан), самое маловероятное событие могло стать почти неизбежным.
РАЗВИТИЕ ЖИЗНИ НА ЗЕМЛЕ (КРАТКИЙ ОЧЕРК)
ДОКЕМБРИЙ
.
Самая древняя эпоха развития жизни -
докембрийская - длилась невероятно долго: свыше
3 млрд. лет.
Выше было рассказано об условиях, в которых жили первые живые организмы. Пищей им служил «первичный бульон» окружающего океана или их менее удачливые собратья. Постепенно, однако, в течение миллионов лет этот бульон становился все более «разбавленным», и, наконец, запасы питательных веществ исчерпались.
Развитие жизни зашло в тупик. Но эволюция благополучно нашла из него выход. Появились первые организмы (бактерии), способные с помощью солнечного света превращать неорганические вещества в органические.
Чтобы строить свои организмы, всему живому требуется, в частности водород. Зеленые растения получают его, расщепляя воду и выделяя кислород. Но бактерии этого делать еще не умеют. Они поглощают не воду, а сероводород, что гораздо проще. При этом выделяется не кислород, а сера. (Поэтому на поверхности некоторых болот можно встретить пленку из серы).
Так и поступали древние бактерии. Но количество сероводорода на Земле было довольно ограничено. Наступил новый кризис в развитии жизни.
Выход из него «нашли» сине-зеленые водоросли. Они научились расщеплять воду. Молекулы воды - непростой «орешек», не так-то легко «растащить» водород и кислород. Это в 7 раз труднее, чем расщепить сероводород. Можно сказать, что сине-зеленые водоросли совершили настоящий подвиг. Это произошло 2 млрд. 300 млн. лет назад.
Теперь в качестве побочного продукта в атмосферу начал выделяться кислород. Накопление кислорода представляло серьезную угрозу для жизни. Начиная с некоторого времени новое самозарождение жизни на Земле стало невозможным - содержание кислорода достигло 1 % от современного. А перед живыми организмами встала новая проблема - как бороться с возрастающим количеством этого агрессивного вещества.
Но эволюция сумела преодолеть и это испытание, одержав новую блестящую победу. Через небольшой промежуток времени на Земле появился первый организм, «вдохнувший» кислород. Так возникло дыхание.
До этого момента живые организмы жили в океане, укрываясь в водной толще от губительных для всего живого потоков солнечного ультрафиолета. Теперь благодаря кислороду в верхних слоях атмосферы возник слой озона, смягчивший излучение. Под защитой озона жизнь смогла выйти на сушу.
Американский писатель-фантаст Клиффорд Саймак в повести «Кто там в толще скал?» так описывает воображаемое путешествие своего героя во времени - в докембрий:
«Дышать было трудно. Кислорода еще хватало,
хоть и с грехом пополам, - из-за этого он и дышал
гораздо чаще обычного. Отступи он в прошлое еще
на миллион лет - кислорода перестало бы хватать.
А отступи еще немного дальше - и свободного
кислорода не оказалось бы совсем.
Всмотревшись в береговую кромку, он приметил, что
она населена множеством крохотных созданий,
снующих туда-сюда, копошащихся в пенном
прибрежном соре или сверлящих булавочные норки в
грязи. Он опустил руку и слегка поскреб камень, на
котором сидел. На камне проступало зеленоватое
пятно - оно тут же отделилось и прилипло к ладони
толстой пленкой, склизкой на ощупь.
Значит, перед ним была первая жизнь, осмелившаяся
выбраться на сушу, - существа, не готовые, да и не
способные оторваться от подола ласковой
матери-воды, которая бессменно пестовала жизнь с
самого ее начала.
Здесь происходило многое, что даст себя знать в
грядущем, но происходило тайно, исподволь.
Снующие козявки и осклизлый налет на скалах -
отважные в своем неразумии предвестники далеких
дней - внушали почтение…»
В течение докембрия природа сделала еще целый ряд замечательных «изобретений». В какой-то момент в клетках образовалось ядро. Примерно тогда же возникло половое размножение, резко ускорившие темпы эволюции. Появились первые многоклеточные существа.
К концу докембрия земные моря населяли разнообразные животные: медузы, плоские черви, губки, полипы. Все они были мягкотелыми, лишенными скелета. Возникновение у животных скелета - раковин, панцирей и т. д. - обозначило начало новой геологической эры.
ЭРА ДРЕВНЕЙ ЖИЗНИ (ПАЛЕОЗОЙСКАЯ).
Палеозойская эра, начавшаяся 570 млн. лет назад, длилась 340 млн. лет. Ученые делят ее на шесть периодов.
Самый ранний из них - кембрий (он продолжался 70 млн. лет). В этот период у самых разнообразных животных начинает развиваться скелет, будь то раковина, панцирь или просто колючие шипики. Видимо, мягкотелость становится к этому моменту слишком небезопасной.
Творчество природы, создающей новые формы жизни, в кембрии необычайно плодотворно и разнообразно: почти все типы животного царства получают своих первых представителей. Хордовых, например, представляют существа, похожие на современного ланцетника. Пропуская воду через жаберные щели, они таким образом процеживают из ила съедобные частички.
Как ни трудно нам представить моря без рыб, но в морях кембрия их еще не было. Моря были густо заселены знаменитыми трилобитами - вымершими предками пауков, скорпионов и клещей.
За кембрием следует ордовик (он длился 60 млн. лет). В море по-прежнему процветают трилобиты. Появляются первые круглоротые - родичи современных миног и миксин. Челюстей у них еще нет, но строение рта позволяет хватать живую добычу, что, конечно, гораздо выгоднее бесконечного процеживания ила.
В следующем периоде - силуре (30 млн. лет) на сушу выходят первые растения (псилофиты), покрывая берега зеленым ковром высотой до 25 см. Вслед за ними на сушу начинают переселятся животные, приучаясь дышать атмосферным воздухом, - многоножки, черви, пауки и скорпионы.
В морях трилобитов уже теснят гигантские ракоскорпионы, длина которых превышает порой 2 м. У позвоночных появляется новый, неизвестный прежде орган - челюсти, развившиеся из безобидных жаберных щелей бесчерепных (например, ланцетника). Чтобы добыча не ускользнула из этих челюстей, рыбы приобретают одновременно парные плавники, увеличивающие маневренность.
Следующий период - девон (60 млн. лет). Сушу заселяют плауны, папоротники, хвощи, мхи. В их зарослях уже живут первые насекомые.
Выбираются на сушу и позвоночные. Как и почему это происходит? Климат в девоне был сухой, температура в течение года резко изменялась. Многие водоемы пересыхали. Некоторые рыбы стали на время засухи зарываться в ил. Для этого нужно было уметь дышать атмосферным воздухом. Но особенно многообещающей для дальнейшей эволюции оказалась группа кистеперых рыб. Помимо легочного дыхания они имели подвижные мускулистые плавники, похожие на лапы. С их помощью они ползали по дну. Чтобы не погибнуть в пересохшем водоеме, кистеперые рыбы отправлялись в сухопутные странствия в поисках воды. При этом они путешествовали на довольно большие расстояния. Естественно, выживали те, которые лучше могли двигаться по суше. Правда, слабых легких для дыхания было недостаточно. Как еще дышать, если жабры на суше не годятся? Только через кожу. Поэтому рыбья чешуя уступила место гладкой влажной коже.
Так в девоне кистеперые рыбы постепенно покинули родную стихию и дали начало первым земноводным - стегоцефалам (панцирноголовым).
Вслед за девоном наступил карбон, или каменноугольный период (65 млн. лет). Впервые огромные пространства суши покрылись болотистыми лесами из древовидных папоротников, хвощей и плаунов.
Глядя на современные небольшие плауны, трудно поверить, что их предки (например, чешуедрев, или лепидодендрон) достигали 40 м в высоту и 6 м в обхвате.
Из падавших в воду и постепенно превращавшихся в уголь стволов образовывались залежи каменного угля. Самый ценный уголь (антрацит) получился из скоплений множества спор, которые роняли в воду деревья того времени.
Сжигая в печке каменный уголь, мы чувствуем тепло солнечных лучей, падавших на Землю без малого треть миллиарда лет назад. Под ними грелись наши далекие предки - земноводные, царствовавшие в карбоне.
Впервые жизнь, освоившая воду и сушу, сделала шаг и в третью стихию - воздух. Первыми и единственными, кто поднялся в воздух в лесах каменноугольного периода, были насекомые. Порой они вырастали до невероятных размеров. Размах крыльев некоторых стрекоз достигал 70 см. А в зарослях помимо пауков и скорпионов стали встречаться, например, тараканы (размером иногда с морскую свинку).
Жизнь сумела окончательно оторваться от породившей ее водной стихии. Почти одновременно это удалось рептилиям и семенным папоротникам, предкам хвойных. У растений появились семена вместо спор, у яиц рептилий - скорлупа. Зародыши в семени и яйце были защищены оболочками, обеспечены пищей. Из яиц рептилий вылуплялся уже не беспомощный головастик, а уменьшенная копия родителя.
Рептилиям уже не нужна была голая кожа для дыхания - вполне хватало легких. Они «заковались обратно в панцирь» из чешуи или роговых щитков.
Последний период эры древней жизни - пермь, или пермский период (55 млн. лет). Климат стал холоднее и суше. Влажные леса из папоротников и плаунов исчезали. Вместо них появились и широко разрослись хвойные.
Земноводных все больше теснили рептилии, шедшие к своему господству на планете.
ЭРА СРЕДНЕЙ ЖИЗНИ (МЕЗОЗОЙСКАЯ).
Мезозойская эра наступила 230 млн. лет назад и длилась 163 млн. лет. Она делится на 3 периода:
* триас (35 млн. лет),
* юру, или юрский период (58 млн. лет), и
* мел, или меловой период (70 млн. лет).
В морях еще в пермский период окончательно вымерли трилобиты. Но это не было закатом морских беспозвоночных. Напротив: на смену каждой форме приходило несколько новых. В течение мезозойской эры океаны Земли изобиловали моллюсками: белемнитами, похожими на кальмаров (их ископаемые раковины зовут «чертовыми пальцами»), и аммонитами. Раковины некоторых аммонитов достигали 3 м в диаметре. Ни у кого больше на нашей планете, ни до того, ни позднее, не было таких колоссальных раковин!
В лесах мезозоя господствовали хвойные, похожие на современные сосны и кипарисы, а также саговники. Мы привыкли видеть насекомых, вьющихся над цветами. Но такое зрелище стало возможным лишь с середины мезозоя, когда на Земле расцвел первый цветок. К меловому периоду цветковые растения уже начали теснить хвойные и саговники.
Мезозой, особенно юру, можно назвать царством рептилий. Но еще в самом начале мезозоя, когда рептилии только шли к своему господству, рядом с ними появились мелкие, покрытые шерстью теплокровные животные - млекопитающие. Долгие 100 млн. лет они жили рядом с динозаврами, почти незаметные на их фоне, терпеливо дожидаясь своего часа.
В юре у динозавров появились и другие теплокровные соперники - первоптицы (археоптериксы). Они имели еще очень много общего с рептилиями: например, челюсти, усеянные острыми зубами. В меловом периоде от них произошли и настоящие птицы.
В конце мелового периода климат на Земле стал холоднее. Природа уже не могла прокормить животных, весивших боле десяти килограммов. (Правда, есть научные теории, иначе объясняющие вымирание динозавров). Началось массовое вымирание (растянувшееся, однако, на миллионы лет) гигантов-динозавров. Теперь освободившееся место могли занять звери и птицы.
ЭРА НОВОЙ ЖИЗНИ (КАЙНОЗОЙСКАЯ).
Кайнозойская эра, начавшаяся 67 млн. лет назад, стала царством птиц, млекопитающих, насекомых и цветковых растений. Она продолжается и сейчас.
Ученые разделяют ее на 3 периода: палеоген,
неоген и антропоген
.
Последний из этих периодов, в котором появляется
человек, начался около 2 млн. лет назад.
ЭВОЛЮЦИОННАЯ ТЕОРИЯ: ДРАМА В БИОЛОГИИ
Эволюционная идея имеет очень древнюю историю: в разной форме мысли о последовательном появлении на Земле живых организмов с постепенно усложняющейся их организацией содержатся и в Книге Бытия, и в «Ригведе», и в произведениях философов древнего Китая и Греции.
Следует заметить, что закономерности эволюционного процесса могут быть выявлены в ходе ключевых проблем. Проблема первая: как возникает многообразие живого - из одного источника (монофилия) или из многих (полифилия) ? Вторая: каков характер возникновения этого многообразия - складывается ли оно медленно и постепенно (градуалистская концепция) или внезапно и быстро (сальтационная концепция) ? Третья: имеют ли эволюционные события случайный, ненаправленный, нецелесообразный характер (тихогенез) или они идут направленно, к определенной цели (моногенез) ?
Ответы Ламарка и Дарвина на первые два вопроса совпадают. Достаточно взглянуть на таблицу Ламарка («Философия зоологии» т. 1 - 2), объясняющую происхождение разных видов животных, чтобы убедиться в его приверженности идеям монофилетизма и дивергентному принципу эволюции).
С этим же принципом тесно связана у Ламарка и Дарвина гипотеза о постепенном, медленном, градуальном характере эволюции.
Но вот в третьем пункте у них есть определенные расхождения.
В отличие от Ламарка, Дарвин допускал участие в эволюции случайных изменений и тем самым отвергал телеологический принцип. Эволюция по Дарвину - не направлена, ее ход непредсказуем. Естественный отбор способен произвести такие формы, появление которых мы можем и не предвидеть.
По Ламарку, организм заранее нацелен на совершенствование и, следовательно, ведет себя более активно. Он - не глина, из которой природа лепит с помощью естественного отбора любые формы, лишь бы они были приспособлены к данным условиям среды. Он, организм, обладает некой собственной внутренней силой, позволяющей ему самому выбирать путь приспособления на основе присущего ему потенциала. Заметим, однако, что этот потенциал не безграничен, он допускает движение только в каких-то ограниченных пределах. То есть эволюция по Ламарку в известной степени телеологична.
Итак, на начальных этапах развития эволюционной теории ламаркизм и дарвинизм фактически не противостояли друг другу. Противостояние возникло позднее. И определила его генетика в ранний период своего становления.
Датский биолог Вильгельм Иогансен писал в начале нашего века, что «генетика вполне устранила основу дарвиновской теории подбора, которая не находит себе никакой поддержки в генетике». Аналогичным образом обстоит дело с гипотезами, которые оперируют такими понятиями, как «наследственные приспособления», «наследование приобретенных свойств» и тому подобными идеями, примыкающими к воззрениям Ламарка. А вот мнение основоположника современной генетики Т. Моргана: «… Естественный отбор не играет созидающей роли в эволюции». Как было упомянуто, еще раньше категорически отверг дарвинизм Грегор Мендель.
Что же заставило этих великих ученых занять столь отрицательную позицию по отношению к дарвинизму и ламаркизму? Прежде всего те открытия, которые вошли в генетику в качестве основополагающих ее постулатов.
Во-первых, генетики выделили два типа изменчивости: модификационную, ненаследственную, и мутационную, наследственную. Наследуются только те признаки, которые возникают в результате изменений в генетическом материале - то есть в результате мутаций. Те же признаки, которые возникают под воздействием условий внешней среды, не передаются потомству. Это - модификации, флюктуации.
Второй, чрезвычайно важный вывод, в корне изменивший взгляды и вошедший в число фундаментальных положений генетики, предельно четко сформулировал Иогансен: «Главным результатом моей маленькой работы является то, что я считаю селекцию флюктуаций совершенно безрезультатной».
Из этих двух заключений логически выводится третье: благоприобретенные признаки не наследуются. Бесчисленные попытки доказать возможность наследования таких признаков потерпели полный крах. Работы Штанфуса, Гаррисона, Тоуэра, Каммерера, Пржибрама, Дюркена в 20-е годы, а Лысенко с сотрудниками в 30-50-е, были экспериментально опровергнуты. В этих работах допускалась одна и та же ошибка: их авторы игнорировали открытия Иогансена и использовали не чистолинейный, а неоднородный в генетическом отношении материал. (Кстати, были опровергнуты и недавние попытки американских иммунологов Горчинского и Стила показать возможность передачи по наследству некоторых приобретенных иммунологических свойств - см. «Nature», 1981, т.259, с.678-681; т.290, с.508-512).
Таким образом, с оформлением генетики как науки отрицание наследования приобретенных признаков было строго экспериментально обосновано и практически общепризнанно. А между тем учение о наследовании приобретенных признаков составляло теоретическую основу представлений и Дарвина, и Ламарка.
Генетики сформировали прямо противоположную концепцию наследственности, в самых общих чертах выдвинутую еще Августом Вейсманом, одним из крупнейших биологов XX века. Он провел резкую грань между сомой - совокупностью клеток, тканей и органов - и клетками зародышевого пути, содержащими зародышевую плазму. В ядрах половых клеток, в их специализированных структурах - хромосомах - собраны гены, носители наследственности, которые передаются от поколения к поколению. Какого-либо переноса частиц от соматических элементов к половым не существует. Таким образом, материальные основы наследственной и ненаследственной изменчивости, общие в теориях Дарвина и Ламарка, в генетике были разделены. Наследственны только те изменения, которые происходят в зародышевой плазме - в генах. Изменения, происходящие в соме, ненаследственны, это - модификации.
Четвертое, важное для судеб эволюционного учения положения генетики, было разработано голландским биологом Гуго де Фризом и русским ботаником °С.И. Коржинским. Вот как это формулировал де Фриз:
1) новые элементарные виды возникают внезапно,
без переходов;
2) новые формы появляются сбоку главного ствола;
3) новые элементарные виды по большей части
вполне постоянны с самого начала своего
возникновения;
4) некоторые из новых форм являются настоящими
элементарными видами, тогда как другие носят
характер ретрогрессивных разновидностей;
5) эти новые формы появляются обыкновенно в
большом числе особей;
6) мутационная изменчивость не связана
непосредственно с модификациями и независима от
них;
7) мутации происходят почти во всех возможных
направлениях;
8) способность к мутации наступает периодически.
И, наконец, пятое принципиальное положение, привнесенное генетикой, - реабилитация понятия вида как такового: вид - не удобная выдумка биологов, а реальная сущность, имеющая достаточно четкие границы и характеризующаяся собственным набором морфофизиологических признаков (фенотипом).
Из изложенного совершенно очевидно, что взгляды родоначальников генетики на движущие факторы эволюционного процесса, в особенности на изменчивость и наследственность, существенно отличны от воззрений Ламарка и Дарвина.
Тем не менее в 30-50-е годы трудами Добжанского, Симпсона, Майра и других была предпринята попытка примирить генетику с дарвинизмом. Так появилась синтетическая теория эволюции (СТЭ), создатели которой стремились обобщить все накопленные генетикой факты и в приложении к эволюционному учению объединить их с позиций Дарвина.
Основа построения синтетической теории эволюции - закон Харди-Вайнберга. Он наглядно показал, что благодаря синтезу с генетикой эволюционное учение постепенно утратило описательный характер и превратилось в математизированную теорию.
Знакомству с законом должно предшествовать краткое вступление о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира.
Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До конца 20-х годов текущего столетия данные генетики противопоставлялись эволюционной теории, наследственная изменчивость (мутационная, комбинативная) рассматривалась в качестве главного фактора эволюции, естественному отбору отводилась второстепенная роль. Таким образом, уже в начальный период своего становления генетика была использована для создания новых концепций эволюции. Сам по себе этот факт знаменателен: он свидетельствовал о тесной связи генетики с эволюционной теорией, но время их объединения было еще впереди. Различного рода критика дарвинизма была широко распространена вплоть до возникновения СТЭ.
Исключительную роль в развитии эволюционного учения сыграла популяционная генетика, исследующая микроэволюционные процессы в природных популяциях. Основана она выдающимися отечественными учеными С.С. Четвериковым и Н.В. Тимофеевым-Ресовским.
Начавшееся в 20-х годах объединение дарвинизма и генетики способствовало расширению и углублению синтеза дарвинизма с другими науками. 30-40-е годы принято считать периодом становления синтетической теории эволюции.
В западных странах обновленный дарвинизм, или синтетическая теория эволюции, приобрел широкое признание среди ученых уже в 40-х годах, хотя всегда были и есть отдельные крупные исследователи, занимающие антидарвиновские позиции.
Основные положения СТЭ выводятся как следствия из закона Харди-Вайнберга. Известно, что понимание сущности и значения закона вызывает у школьников затруднение, хотя его математический аппарат прост и доступен всем, кто знаком с алгеброй средней школы. Важно сосредоточить внимание учащихся не только на определении закона - частоты генов и генотипов в популяции не меняются в ряду поколений, - его условиях - бесконечно большая популяция, случайное свободное скрещивание особей, отсутствие мутационного процесса, естественного отбора и других факторов, - математической модели - AA p2 + Aa 2 p + aaq2 = 1, - но и на практическом применении закона.
Процесс микроэволюции доступен непосредственному изучению, о его наличии можно судить по изменяющемуся генофонду. Генофонд описывается либо частотами аллелей, либо частотами генотипов, соотношение которых легко установить. Для этого нужно подсчитать гомозиготных носителей рецессивного признака и, пользуясь простой формулой, вычислить частоту встречаемости аллелей, доминантных гомозигот и гетерозигот. Зная соотношение генотипов и аллелей в данной популяции, можно проследить ее дальнейшую эволюционную судьбу. Например, один из 10000 человек альбинос, т.е. частота гомозигот по рецессивному аллелю (q2) составляет 0,0001. Зная, что q2 = 0, 0001, можно определить частоту аллеля альбинизма (q), доминантного аллеля нормальной пигментации (p), гомозиготного доминантного генотипа (p2), гетерозиготного генотипа (2 pq) :
p2 = 0,01 (или 1 %).
Поскольку p = 1 - q, то 1 - 0, 01 = 0,99; частота доминантного аллеля в популяции равна 0,99, или 99 %. Нормально пигментированным людям будут соответствовать два генотипа (АА, Аа). Гомозиготный доминантный генотип будет встречаться с частотой 0,98, так как p2 = 0,992 = 0,98. Частота встречаемости гетерозигот составляет примерно 0,02, или 2 %, так как 2pq = 2 x 0, 99 х х 0,01 = 0,0198 = 0,02.
Важно обратить внимание на следствия, вытекающие из закона:
* рецессивные аллели присутствуют в популяции
главным образом в гетерозиготном, а не в
гомозиготном. Состоянии. (Как видно из
рассмотренного примера, при малом числе
альбиносов, 200 человек из 10000 (2 %) несут аллель
альбинизма в скрытом, гетерозиготном состоянии);
* в отличие от идеальной популяции на популяцию в
природе оказывают давление эволюционные
факторы, которые приводят к изменению ее
генофонда.
В процессе дальнейшего изложения материала следствия закона Харди-Вайнберга будут углубляться, конкретизироваться. Так, при рассмотрении роли изменчивости в эволюционном процессе, что именно исходя из закона Харди-Вайнберга отечественный генетик С.С. Четвериков первым высказал мысль о значении рецессивных мутаций: они не исчезают в популяции, не растворяются в массе нормальных особей, а поглощаются свободным скрещиванием, оставаясь скрытыми в гетерозиготном состоянии в недрах вида, и проявляются фенотипически только в популяции. С работы С.С Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926) и начался период синтеза генетики и эволюционного учения. Ученый сделал важный вывод о насыщенности природных популяций большим количеством рецессивных мутаций. Существование такого скрытого резерва наследственной изменчивости создает возможность для эволюционных преобразований в популяции под воздействием естественного отбора.
Таким образом, мы подводим к пониманию положений синтетической теории эволюции о том, что микроэволюционные процессы начинаются в популяции; материалом для эволюции служат чаще всего рецессивные мутации.
Другое следствие закона раскрывается при изучении эволюционных факторов. Условия идеальной популяции не соблюдаются в природе. Идеальная популяция не тождественна реальной. Под давлением эволюционных факторов частоты аллелей и генотипов в популяции изменяются, и это является основой всех эволюционных преобразований. Понятие каждого конкретного фактора эволюции вводится как результат поочередной замены условий идеальной популяции на противоположные. Например, если постоянство частот аллелей и генотипов в идеальной популяции имеет силу при бесконечно большой численности особей, то в малой природной популяции случайные процессы могут привести к заметным последствиям, в частности, к изменению ее генофонда. Такое противопоставление позволяет логично перейти к рассмотрению дрейфа генов - ненаправленного случайного изменения частот аллелей в малых популяциях, одна из причин которого - популяционные волны.
И еще. В идеальной популяции все генотипы одинаково плодовиты, т.е. естественного отбора не происходит, на реальную же популяцию естественный отбор оказывает существенное давление. В связи с этим раскрывается сущность естественного отбора как направляющего эволюционного фактора, выявляются его формы, творческая роль.
Рассмотрение факторов эволюции на основе закона Харди-Вайнберга позволяет перейти к объяснению ее результатов: приспособленности и видообразования. Раскрывая способы видообразования, важно отметить, что этот процесс может быть постепенным (географическое и экологическое видообразование) и внезапным. Внезапное видообразование одно время противопоставлялось его дарвиновскому пониманию. Современная эволюционная биология допускает кроме постепенной дивергентной эволюции внезапное видообразование путем крупных хромосомных мутаций, полиплоидии, отдаленной гибридизации. При этом сущность процесса видообразования одинакова для всех способов и заключается в коренной перестройке свойств родительского вида и формировании свойств нового вида.
В заключении сформулируем основные положения синтетической теории эволюции, дополнив их идеями современной эволюционной биологии, не вошедшими в синтетическую теорию эволюции.
ОСНОВНЫЕ ПОЛОЖЕНИЯ СОВРЕМЕННОЙ ТЕОРИИ ЭВОЛЮЦИИ:
Наименьшая эволюционная единица, в которой проходят микроэволюционные процессы, - популяция.
Материалом для естественного отбора служат, как правило, очень мелкие, дискретные единицы наследственности - мутации.
На популяцию оказывают давление эволюционные факторы: мутационный процесс, волны численности, изоляция, естественный отбор.
Мутационный процесс, волны численности - факторы-поставщики материала для эволюции - носят случайный и ненаправленный характер.
Изоляция усиливает действие факторов-поставщиков эволюционного материала, ее давление на популяцию также не направленно.
Единственный направляющий фактор эволюции - естественный отбор.
Эволюция носит постепенный, дивергентный и длительный характер, этапом которой является видообразование.
Вид состоит из множества подвидов и популяций.
Обмен аллелями; «поток» генов возможны лишь внутри вида; вид - генетически целостная и замкнутая система.
Эволюция непредсказуема.
Важно подчеркнуть, что синтетическую теорию эволюции не следует рассматривать как законченное учение. Нужно внести дополнения и поправки, выдвинутые современной эволюционной биологией:
В небольших изолированных популяциях важный фактор эволюции - дрейф генов.
Эволюция носит не только постепенный, но и внезапный характер.
Эволюция может быть предсказуема. Оценивая прошлую историю, генотипическое окружение и возможное влияние среды, можно предсказать общее направление эволюции.
Изучение эволюционной теории на основе опорных знаний по дарвинизму и генетике позволяет показать эволюционную биологию как интенсивно развивающуюся науку, где широко используются математические закономерности.
Однако, синтетическая теория, при всей ее заманчивости, вряд ли может быть признана единственно возможной основой эволюционного учения. И в связи с этим следует отметить концепцию номогенеза Л.С. Берга (Избранные труды. М.: «Наука», 1977), постулаты которой альтернативны дарвинизму.
В концепции номогенеза большое значение придается скачкообразным, внезапным изменениям. Основу таких изменений могут составлять системные мутации, которые коренным образом преобразуют процессы индивидуального развития и приводят к появлению так называемых «счастливых монстров» или «перспективных уродов» - организмов, существенно отличающихся от родительских.
М.Д. Голубовский, основываясь на открытии генетических подвижных элементов и многочисленных данных генетико-популяционного анализа, предложил новую триаду факторов эволюционного процесса (взамен неодарвинистской: наследственность - изменчивость - отбор). Это: облигатный геном (ОГ) - факультативный геном (ФГ) - фактором среды (Молекулярные механизмы генетических процессов. М.: «Наука», 1985, с.146-162).
ПО ЧАРЛЬЗУ ДАРВИНУ:
1. Организмы развились из одной или немногих
первичных форм.
2. Развитие шло на основе случайных вариаций
отдельных особей.
3. Развитие шло путем медленных небольших
изменений.
4. Наследственных вариаций много, и они идут по
всем направлениям.
5. Виды связаны друг с другом постепенными
переходными формами.
6. Эволюция состоит в образовании новых
признаков.
7. Борьба за существование и естественный отбор
являются факторами прогресса (эволюции).
ПО Л.С. БЕРГУ:
1. Организмы развивались из многих тысяч
первичных форм.
2. Развитие шло на основе закономерностей,
захватывающих массу особей.
3. Развитие шло скачками, параксизмами.
4. Число наследственных вариаций ограничено, и
они идут по определенным направлениям.
5. Разные виды резко разграничены в силу
скачкообразного происхождения.
6. Эволюция состоит в значительной степени в
развертывании задатков.
7. Борьба за существование и естественный отбор -
консервативный фактор, охраняющий норму.
Согласно этой гипотезе геном подразделяется на две части: облигатную, или постоянную (эквивалент наследственности в классической триаде), обеспечивающую консерватизм вида, и факультативную (эквивалент наследственной изменчивости), которая представляет собой совокупность подвижных генов. Они придают геному пластичность, то есть его направленное преобразование, и тем самым определяет возможность эволюции. Фактор, активирующий изменения ФГ, - внешняя среда, хотя характер ее воздействий на геном пока не ясен.
Однако, представляется, что условия активности ФГ - не вне живой системы, а присущи ей. Иными словами, это не только (а может быть, не столько) внешняя среда. В качестве эволюционно значимой части генома правильнее рассмотреть, помимо ФГ, то есть совокупности генетически подвижных элементов, еще и те структуры генома, которым избирательно «сродственны» эти элементы. Ведь известно, что места их фиксации в геноме зачастую не случайны. Вероятно, подвижные гены активируют преобразования именно в тех частях генома, которые изначально предназначены для реализации этих процессов. Если это так, то план эволюционного развития фиксирован в геноме точно так же, как и план индивидуального развития, и осуществляется этот план благодаря специфическому взаимодействию подвижных элементов с определенной, предназначенной для этой цели частью генома (назовем ее «программирующим геномом» - ПГ).
Следовательно, взаимодействие типа ФГ+ПГ способно вызвать к жизни программированный, но не проявленный до того морфогенез, ибо, тот или иной морфогенез может быть не осуществлен не потому, что эволюционно не сформировались его генетическая программа, а потому, что в генетическом материале содержатся элементы, тормозящие проявление этой программы. Удаление подобных элементов (а также их перераспределение в геноме) существенно преобразовывает функциональную организацию генома в целом, так что открываются новые морфогенетические пути. На основе измененного типа онтогенеза возникают организмы с новыми фенотипическими признаками, которые можно считать соответствующими «многообещающим монстрам»…
Таким образом, ОГ обеспечивает полиморфизм, разнообразие, но лишь в пределах данного вида, то есть изменения в ОГ не затрагивают набора видоспецифических характеристик. А вот взаимодействия ПГ+ФГ вызывают эффективные изменения уже не отдельных признаков, а генома в целом, переводя его на новый структурно-функциональный уровень и выводя за пределы данного вида. Так появляются новые виды. Отсюда правильнее подразделять геном не на два, а ни три части: облигатную, факультативную и программирующую.
Эти три относительно автономные части генома составляют в то же время неразрывное единство, и функция генома в целом определена взаимодействием всех его компонентов.
Итак, можно подвести итог, сделав тривиальное заключение: современное состояние эволюционного учения отнюдь не простое, и наряду с ортодоксальным направлением, неодарвинистским, «синтетическим», существуют взгляды, так или иначе им противоречащие. В целом это хорошо. Время догматизма в эволюционной биологии себя исчерпало. В конце концов, наблюдаемые сегодня очевидные успехи молекулярной генетики, генетики развития, равно как и становление серьезных социобиологических подходов, создают реальные предпосылки к постепенному ограничению числа эволюционных гипотез - теперь уже на строго научной основе. А это, хочется верить, означает, что до истины не так далеко. И у эволюционизма - этой перманентной драмы биологии - все-таки будет всех примиряющий финал.
ПРИГОВОР… ИЗ БИБЛИИ
В тот древний, исступленный век,
Когда Творец хулил свое творение
И созданное рушил…
Рабиндранат Тагор
История растительного и животного мира полна примеров возникновения, расцвета и гибели отдельных видов, родов и даже целых отрядов.
Первое массовое вымирание целой группы организмов произошло в конце кембрия, когда исчез загадочный тип археоциат. Он существовал менее 100 млн. лет и является единственным типом животных с таким коротким временем существования - все другие имеют возраст более 500 млн. лет. В конце кембрийского периода вымирает и часть трилобитов, но группа в целом сохраняет свою стабильность за счет появления новых родов. Граница девона и карбона (350 млн. лет назад) является началом одного из глобальных кризисов жизни. Вымирают псилофиты - завоеватели суши, практически вымирают граптолиты, начинается гибельный упадок трилобитов и гигантских ракоскорпионов, резко уменьшается число наутилоидей. Этот глубокий кризис продолжается около 100 млн. лет, до конца палеозойской эры (конца перми).
В конце палеозойской эры жизнь подводит первые «итоги» этого великого перелома. Вымирают полностью: трилобиты, гигантские ракоскорпионы, формаминиферы семейства фузулинил, древние иглокожие (бластоидеи, древние морские ежи, древние морские лилии), четырехлучевые кораллы и их родственники табулята, крупные группы брахиопод, большинство наутилоидей, древние аммоноидеи. Резко сократились древние группы земноводных и некоторые древние пресмыкающиеся. Полностью исчезают гигантские лепидодендроны, сигиллярии, каламиты, многие папоротники. Итак, граница палеозоя и мезозоя отмечена массовым исчезновением крупных групп организмов и появлением новых. На месте исчезнувших групп развивается богатая и разнообразная голосеменная растительность (гинковые, цикадовые, хвойные), белемниты и аммониты со сложными складчатыми перегородками, многочисленные группы пресмыкающихся и другие.
Второй глобальный кризис в течение фанерозоя произошел в конце мезозойской эры. Массовое вымирание захватывает большие группы пресмыкающихся (динозавров, летающих ящеров, ихтиозавров и плезиозавров), вымирают аммониты и делемниты, толстозубые двустворчатые моллюски пахиодонты, иноцермы, большая группа цветковых растений и другие. 2-3 млн. лет назад исчезает ряд гигантских млекопитающих».
«Для объяснения массового вымирания больших групп организмов создано так много гипотез, - пишет Т.Г. Николаев в труде «Долгий путь жизни», - что только их критическому рассмотрению известный советский палеонтолог Л.Ш. Давиташвили посвятил книгу объемом более двухсот страниц. Почти все, кто касался этой проблемы, - от авторитетных ученых до бойких популяризаторов, - все считали необходимым создать собственную теорию.
Большинство гипотез настолько наивны, что только ради этого можно их почитать как развлекательную книгу в часы досуга. Разумеется, существует обоснованная научная теория глобальных кризисов в истории жизни - это учение Дарвина, развившаяся позже и представленное сегодня современной синтетической теорией эволюции».
Зададимся вопросом: объясняет или не объясняет это «учение» Тайну Глобальных Кризисов…
По Дарвину, «вымирание и естественный отбор идут рука об руку». «Теория естественного отбора основывается на том положении, что каждая новая разновидность и в конце концов каждый новый вид возникает и держится благодаря тому, что он имеет некоторое преимущество над теми, что он имеет некоторое преимущество над теми, с которыми ему приходится конкурировать; из этого почти неизбежно следует вымирание форм менее благоприятствуемых».
Таким образом, по Дарвину, исчезновение огромного количества прежних форм связано с тем, что они были вытеснены, истреблены, побеждены в конкурентной борьбе новыми, более совершенными и более приспособленными формами. Но большинство современных ученых, писавших по вопросу о вымирании организмов, считают дарвинистское объяснение этого процесса устаревшим. Эти ученые еще соглашаются допустить, что биоценотические факторы играли некоторую роль в отдельных частных случаях вымирания небольших групп, если эти группы не имели очень широкого распространения. Но обычно считается невозможным привлекать биоценотические факторы к объяснению вымирания обширных групп животных и растений, господствовавших в морях или на суше в прежние геологические периоды. В смене господствующих групп наблюдается некоторая закономерность, не совпадающая с той, какая рисовалась ортодоксальными последствиями дарвинистской теории. При рассмотрении фаун отдельных периодов видны, напротив, многочисленные примеры другого рода, противоречащие представлению о конкуренции как о постепенном вытеснении одних групп другими, более приспособленными. На самом деле чаще видно как раз обратное, именно предшествующее вымирание и наличие свободных мест в экономии природы является толчком для развития новых групп.
Дарвиновское понимание вымирания Депере (1921) называет «остроумным объяснением, построенным на борьбе за существование». «Так как непосредственная борьба с другими видами казалась неприложимой к крупным млекопитающим - исполинским динозаврам, то знаменитый преобразователь эволюционного учения обращается к затруднительности для этих гигантских животных находить себе достаточное количество пищи - объяснение почти детски наивное, так как оно касается травоядных животных, населявших такие беспредельные материки, какими должны были быть обширные юрские равнины в центре и на востоке теперешних Соединенных Штатов». «Как может «адаптация» (понимаемая как выработка более целесообразного в борьбе за существование) вести к вымиранию?» - справедливо спрашивает А.А. Любищев (1946).
Все это привело к тому, что подавляющее большинство ученых, работающих над проблемой вымирания, приходят обычно к выводу, что концепции Дарвина недостаточны, неудачны, наивны и не соответствуют нынешнему уровню биологических знаний. Короче говоря, современные ученые считают целесообразными при изучении проблемы вымирания обходиться без Дарвина. Так, Соболев пишет (1924) : «У нас нет никаких положительных оснований для утверждения, что в истреблении трилобитов, например, повинны головоногие… Точно так же вымирание древних наутилоидей невозможно поставить в связь с распространением в готландии высоко развитых рыб, и не только потому, что своей хорошо развитой раковиной едва ли представляли подходящий корм для рыб, но и потому еще, что при таком допущении было бы совершенно непонятно, почему такого истребления избежали только что появившиеся аммоноидеи (нониатиды).
Еще менее понятно, почему палеозойские рыбы не могли справиться со сравнительно мелкими и слабыми гониатидами, а приобретшие широкое распространение в мелу костистые рыбы уничтожили, как полагает Неймайр, аммонитов и белемнитов.
Столь же проблематичным представляется непосредственное вытеснение стегоцефалов рептилиями, и уже труднее указать, где те более приспособленные, которые сжили со света роскошную фауну мезозойских рептилий. Мы не можем сказать, что это были млекопитающие, ибо те из них, которые жили в эру господства рептилий, были слишком ничтожны и малочисленны, чтобы вести с ними борьбу, даже хотя бы, как это некоторые полагают, путем поедания их яиц, а более высоко организованные и многочисленные плацентарные млекопитающие не вытеснили рептилий, а заняли место уже ставшее свободным после их вымирания.
Все это, равно как и одновременное вымирание в определенные моменты времени сразу многих и часто весьма разнообразных форм, трудно объяснить одной борьбой за существование».
Что же касается климатических, геологических и других подобных причин, то «… многие ученые справедливо отмечают, что изменениями климата нельзя объяснить повсеместное вымирание тех или иных групп в определенные геологические эпохи. А.П. Павлов (1924) писал об этом вполне справедливо: «Конечно, климатические изменения имели большое влияние на судьбу органического населения какого-нибудь древнего континента или его части. Но ведь климаты Земли не изменялись внезапно и повсюду одинаково…»
Прав и Э. Берри (1923), который, говоря о причинах вымирания животных, признает влияние изменений климата и других условий среды, но и в то же время сомневается в том, что эти изменения, которые совершались не внезапно, имели когда-либо значение первостепенных факторов.
В поисках причин вымирания многие геологи уделяют особое внимание изменениям в распределении суши и моря, в конфигурации материков и океанов, но никакие физико-геологические изменения не могли вызвать исчезновение больших групп наземных и водных организмов, которые до того были распространены по всему земному шару.
Некоторые ученые склонны думать, что тектонические «перевороты» представляют собой главную, или даже единственную, непосредственную причину вымирания. Однажды против этого возражал Вальтер, указывая, что даже поднятие Альп и Гималаев происходило так медленно, что это вызывало медленную миграцию флоры и фауны этих складчатых полос, но не наносило ущерба населению.
Таким образом, в современной палеонтологии происходит «…весьма широкое распространение пессимистической оценки возможности выяснить причины, вызывающие вымирание животных и растений». Так, известный германский палеонтолог и геолог К. Динер (1920) говорит, что «совершенно темной останется причина вымирания аммонитов и динозавров в конце мелового времени и гибель Gigantostracca и трилобитов с начала девона, спираленосных плеченогих в триасе, причина внезапного появления и угасания больших по всему миру распространенных групп фораминифер». Далее он говорит: «Попытка объяснить неоднократное внезапное исчезновение больших групп, не оставляющих потомства, встречает пока едва ли преодолимые затруднения». Динер отмечает тот поразительный факт, что один из наиболее значимых переломов в истории наземных позвоночных, совершившийся на грани мелового периода и палеогена, не сопровождался соответствующим ему изменением в характере мира растений. Этот, по его словам, важный и удивительный факт делает еще более непонятной смену крупных рептилий верхнего мела малорослыми млекопитающими палеоцена.
Говоря о всесветном вымирании гигантских рептилий моря и суши в конце мелового периода, Осборн (1910) утверждал, что человечество не имеет никакого понятия о том, какой всесветной причиной можно объяснить это явление. Пресмыкающиеся чувствительны к температуре, и было бы естественно приписать их вымирание общему понижению температуры, но ископаемые флоры Европы и Америки, по его словам, не указывают на это. Нет также признаков какого-либо большого географического катаклизма, и растительность мела в области Скалистых гор постепенно переходит в нижнетретичную.
Массовые вымирания давно волнуют биологов. Шутка ли, когда в считанные миллионы лет гибнут пучки, представленные тысячами филетических линий. Достаточно вспомнить о гибели наиболее продвинутых по пути прогресса рептилий в конце мелового периода, причем таких разных, как динозавры на суше, птерозавры в воздухе и ихтиозавры в воде. В конце концов гибель всегда можно как-то объяснить (взрыв сверхновой звезды, падение астероида, массовые извержения вулканов и прочее) ; непонятно только, почему одни гибнут, а другие, вроде бы не лучше и не хуже, продолжают процветать.
Вот мы и пришли к выводу: никакие экологические и тому подобные факторы не являются причиной гибели древних организмов, так как «история мира организмов показывает, что массовые вымирания касались не экологических групп, а систематических (например, вымирают все динозавры - морские, наземные, летающие) ». Систематически же, избирательно и целенаправленно может действовать только разум, который уничтожал одни систематические группы как не понравившиеся Ему и оставлял в живых другие, более красивые и совершенные.
Эти виды были просто уничтожены людьми, Высшими Людьми - Богами, так как их форма и строение не отвечали законам Красоты и Гармонии. Эти виды не удовлетворяли Их своим несовершенством, и поэтому Они эти виды безжалостно уничтожили. Так были уничтожены панцирные рыбы, аммоноидеи, трилобиты, стегоцефалы, саблезубые тигры, мастодонты и динозавры.
Да, да! Динозавры не вымерли. Они были уничтожены как тупиковая ветвь, как неудавшиеся формы творчества, как чисто экспериментальные образцы.
«…Завершается меловой период, а с ним и вся мезозойская эра. И происходит невероятное - динозавры исчезают! Колоссы и пигмеи, сухопутные, морские и летающие. Все до одного, не оставив потомства! Из 16 огромных отрядов рептилий, населявших Землю в мезозое, до наших дней дожили лишь 5: крокодилы, ящерицы, черепахи и гаттерия.
Вымирание динозавров произошло сразу на всех материках и во всех природных зонах. Особенно загадочно исчезновение морских рептилий. У них не было природных врагов - водные млекопитающие появились намного позже, перейдя с суши в океан, уже «очищенный» к тому времени от динозавров.
Морские рептилии были весьма мобильны, питались в основном рыбой, многие из них (скажем ихтиозавры) были живородящими. Все это делало их весьма защищенными от любых климатических, геологических или экологических катастроф. «Великое вымирание» динозавров произошло, конечно, не за один день, но все же по геологическим меркам почти мгновенно. Известны так называемые поля смерти динозавров в Средней Азии. На некоторых участках вдоль подножий Тянь-Шаня тянутся гигантские скопления этих ящеров - останки миллионов и миллионов особей. И.А. Ефремов, руководивший раскопками в Монголии, отмечал, что нередко в захоронениях одновременно встречаются скелеты старых животных и молодняка. Это могло произойти только вследствие какой-то внезапной катастрофы (или внезапного целенаправленного действия). С исчезновением динозавров млекопитающие не просто обрели «жизненное пространство». Произошла мощная вспышка эволюционного развития этого класса животных».
Динозавры исчезли примерно 65 миллионов лет назад. «Ни те, кто приспособился к бегу на суше, ни плавающие в лагунах, ни легко планирующие на крыльях, ни закованные в роговые панцири тихоходы - никто не уцелел.
Погибли плотоядные и растительноядные. Ни одна из веточек этого широкого развитого рода не смогла перебраться через границу, отделяющие мезозойскую эру от кайнозоя - среднее время от нового», - пишется в одной популярной статье.
Но почти так же описывает эти события и один из корифеев палеонтологии Д. Симпсон (1948) : «Самое загадочное событие в истории Земли - это переход от мезозоя, века рептилий, к кайнозою, веку млекопитающих. Впечатление такое, словно во время спектакля, в котором все главные роли играли рептилии, и в частности толпы самых разнообразных динозавров, занавес на мгновение упал и тотчас взвился вновь, открыв те же декорации, но совершенно новых актеров: ни одного динозавра, прочие рептилии на заднем плане в качестве статистов, а в главных ролях - млекопитающие, о которых в предыдущих действиях и речи не было».
К этому надо добавить, что почти во всех статьях, посвященных внезапной гибели динозавров, как-то все время упускается из виду, что в это же самое время происходила массовая и внезапная гибель других организмов - одновременно с динозаврами исчезли тогдашние черепахи, рыбы, моллюски, растения…
О гибели же динозавров в конце мелового периода говорят чаще всего и больше всего, потому что они наиболее заметны и их гибель бросается в глаза в первую очередь. Таким образом, в конце мела происходила не просто гибель динозавров, а внезапное исчезновение целых фаун и флор.
Г. Мартисон (1980), проводивший раскопки в пустыне Гоби, пишет: «Нас поразило огромное скопление ископаемых рыб, заключенных в большие плиты плотных песчаников. Встречались также обломки костей динозавров и черепах, целые горизонты с раковинами моллюсков и остракод.
В отложениях прибрежных участков собраны остатки различных насекомых. В одних горизонтах встречались отпечатки папоротников и листья иных растений, в других - раковины моллюсков, скелетные остатки ракообразных, рыб и множество насекомых. Большое количество остатков стволов ископаемых болотных кипарисов… Неоднократно попадались кости динозавров и целые горизонты с панцирями черепах. Эти панцири в некоторых местах лежали так густо, что напоминали булыжную мостовую. Невольно напрашивается вопрос: какая причина могла вызвать столь массовую гибель этих животных?»
Динозавры погибли на всех материках и во всех природных зонах. Погибли маленькие и большие, наземные и водные, хищные и травоядные, слабые и те, у которых не было врагов. Погибли старые и молодые, масса отложенных яиц, из которых никто не вылупился.
Смерть была внезапной, что доказывают «находки больших скоплений крупномерных раковин моллюсков, погибших еще в прижизненном состоянии, о чем свидетельствует плотная сомкнутость створок большинства особей» (Г. Мартинсон, 1980). Она была мгновенной, как удар молнии. Так, в «Улан-Баторе в палеонтологической лаборатории геологического института АН МНР выставлены скелеты двух динозавров, сцепившихся в мертвой схватке. Небольшой хищный динозавр схватил в свои объятия рогатого динозавра - протоцератопса и вместе с ним был захоронен на илистом дне водоема» (Г. Мартинсон, 1980). Подобный экспонат мог появиться только в том случае, если смерть одновременно настигнет их обоих, если бы они упали в водоем (в болото) еще живыми, то барахтаясь в воде обязательно расцепились бы.
М. И. Голенкин (1927) говорит про «колоссальное изменение животного мира к концу мелового периода», о «революции на границе нижнего и среднего мела, с геологической точки зрения протекавшей как бы в период покоя Земли». Он думает, что всю доступную нам историю Земли и ее обитателей можно разделить на два периода: до появления покрытосеменных и после их появления. На границе нижнего и среднего мела произошла, по его мнению, своеобразная катастрофа, не связанная ни с извержением вулканов, ни с землетрясением, но отразившаяся глубочайшим образом на дальнейшем существовании всех живых существ.
Итак, 65 миллионов лет назад полностью исчезли исключительно распространенные в мезозое рудисты, аммоноидеи, наземные, морские и летающие ящеры. Это значит, что «некто в сером» взял и сразу выключил все это множество биологических игрушек, выставленных в витрине магазина под названием «Природа».
Все сводится к тому, что динозавры погибли не потому, что началось широкое распространение млекопитающих, как это считает академик Л. Габуния, ибо млекопитающие в пору динозавров были жалкими существами не более крысы; кроме того, как указывает Д. Симпсон (1948), млекопитающие распространились лишь после исчезновения динозавров. Динозавры погибли не от похолодания, не от отравления, не от эпидемии, не от усиления космической радиации, не от кислородного голодания, не от изменения пищевых ресурсов, не от перемены магнитного поля Земли, не от катастрофы, связанной с падением астероида или взрывом близнаходящейся звезды… Ибо никакая катастрофа не приводит к определенно-выборочному характеру «вымирания». Никакая катастрофа не уничтожает сильных, более защищенных, от которых нечего ждать в будущем, для того чтобы дать жизненное пространство и свободу менее защищенным, еще слабым и маленьким, но более высокоорганизованным. Ни у какой катастрофы не может быть такой верной мысли, такой железной логики и таких далеко идущих планов. Эта мысль, эта логика, эти планы были у Тех, кто создал жизнь на нашей планете, кто развивал ее в новые виды. Это не катастрофа, а сознательный акт массового уничтожения Мастером не понравившихся Ему творений. «И вот Я истреблю их с Земли», - сказано в Библии (Бытие, глава 6). И Он истребил их с Земли. Провозившись с динозаврами долгое время, создав их великое множество - самых разнообразных, но найдя их всех несовершенными, Мастер безжалостно уничтожил древних рептилий, оставив в живых только крокодилов, черепах, ящериц и гаттерию (и, возможно, еще какие-то малочисленные роды, скрывающиеся где-нибудь в глубинах океана или в непроходимых джунглях) как наиболее совершенных с художественной и технической точки зрения.
Так же, как гончар безжалостно разбивает неудавшийся кувшин, Высший Гончар безжалостно уничтожил неудавшиеся формы динозавров как не отвечающие законам Красоты и Гармонии. По этой причине были уничтожены и все другие организмы мелового периода. После уничтожения динозавров эти Высшие люди на освободившихся пространствах начали развивать млекопитающих и птиц как более прогрессивные и художественно-совершенные группы организмов.
… И Люциферовы Крыла
Уродства ваши уничтожат, -
Чтоб Красота одна цвела,
Ее уничтоженьем множат!
А. П. Павлов (1924) говорит, что «вымирание в эти эпохи не ограничивалось одними наземными или одними морскими организмами, а захватило тех и других. Как будто это явление обусловливалось одной общей причиной и для разнородных групп, и для существенно разных местообитаний».
Да не как будто, а в буквальном смысле, в действительности это явление обусловливалось одной общей причиной. И такой общей единой причиной является Разум. Высший Разум Высшего художника, уничтожившего не понравившиеся Ему творения.
«Говоря о макроэволюции, мы пользуемся широко принятой аналогией между филетической группой и деревом. Развивая ее дальше, можно сравнивать филетическое видообразование с ростом ветвей. Правда, в отличие от обычного дерева за этим филогенетическим древом следит «садовник», который время от времени подстригает побеги, лишая ветви дальнейшего роста. Этот садовник в своей работе придерживается некоторых правил: во-первых, он подстригает только ветви, расположенные на максимальной высоте, и, во-вторых, этой операции нередко подвергаются все побеги одной крупной ветви, включающей в себя множество более мелких ветвей и веточек». (В.А. Бердников, 1991).
СПИСОК ЛИТЕРАТУРЫ:
1. Сухорукова, «Изучение современной эволюционной теории в X-XI классах».
2. Войткевич Г.В., «Развитие жизни на земле (краткий очерк».
3. «Химия и жизнь», 1993, №1, Шрейдер Ю., «Эволюция и сотворение мира».
4. «Химия и жизнь», 1993, №12, Корочкин, «Эволюционная теория: драма в биологии».
5. «Химия и жизнь», 1997, №1, Травин А., «Этюды по теории и практике. Отбор».
6. «Свет», 1997, №3, Пронин А.В., «Приговор из библии».
7. «Химия и жизнь», 1997, №3, Травин А., «Этюды по теории и практике эволюции. Гены добрые, гены злые».
8. «Студенческий меридиан», 1997, №4, «Рост - двигатель эволюции».
9. «Энциклопедия для детей», 1994, Издание №2,Изд-во «Аванта+».
10. Татаринов, «Суждение палеонтолога об эволюции».
Раздел: «Развитие органического мира» (6 часов)
Эволюционное развитие органического мира.
I. Главные пути и направления эволюции органического мира. Макроэволюция, её доказательства.
Понятие макроэволюции.
Понятие биологического регресса.
Понятие биологического прогресса.
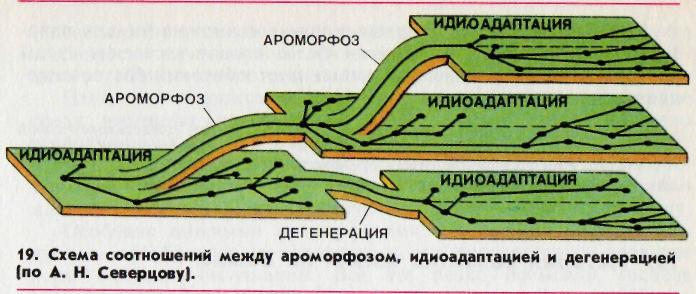
Главные направления биологического прогресса: ароморфоз , идиоадаптация, дегенерация.
Доказательства эволюции: эмбриологические, палеонтологические, сравнительно–анатомические, биохимические, биогеографические.
Понятие макроэволюции.
Понятие биологического регресса.
3. Понятие биологического прогресса.
Биологический прогресс
– направление эволюции, которое приводит к увеличению численности вида, расширению его ареала, увеличению числа популяций вида и образованию новых популяций вида. Биологический прогресс достигается тремя путями: ароморфозом, идиоадаптацией и дегенерацией.
Главные направления биологического прогресса:
Ароморфоз
(арогенез, морфофизиологический прогресс) – эволюционные изменения, которые ведут к общему подъему организации, повышают интенсивность жизнедеятельности, но не являются узкими приспособлениями к резко ограниченным условиям существования.
Ароморфоз дает значительные преимущества в борьбе за существование и делает возможным переход в новую среду обитания, способствует повышению выживаемости и снижению смертности в популяции. Так, усложнение строения древних земноводных по сравнению с рыбами способствовало вступлению земноводных на путь биологического прогресса. Например, у млекопитающих сердце разделено на две половины – правую и левую с развитыми двумя кругами кровообращения. Дифференцировались органы пищеварения.
Идиоадаптация
(аллогенез) – мелкие эволюционные изменения, которые способствуют приспособлению к определенным условиям среды обитания (частные приспособления).
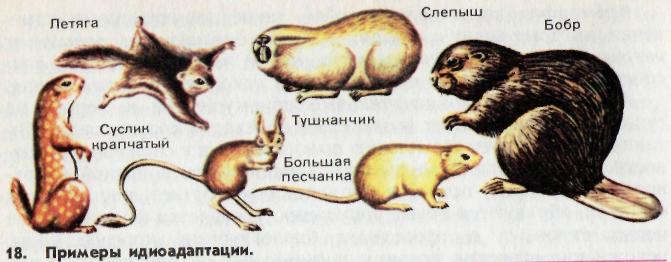
Идиоадаптация не сопровождается изменениями основных черт организации, общим подъемом её уровня и интенсивности жизнедеятельности организмов. Например, в засушливые и жаркие периоды у некоторых земноводных на основе наследственных изменений в процессе борьбы за существование и естественного отбора выработались частные приспособления к определенным и ограниченным местообитаниям – в болотах и топях. Никакого повышения уровня организации у земноводных при этом не произошло. Современные амфибии – лягушки, тритоны – хорошо приспособлены к условиям существования в мелких водоемах и сильно увлажненных местах суши. Приспособления придонных рыб – камбаловых, скатов к жизни на дне (уплощение тела, окраска под цвет грунта) представляют собой типичные примеры идиоадаптации.
Защитная окраска дает хороший пример приспособления животных к определенным условиям существования, не связанного с повышением организации.
Примерами идиоадаптации у растений могут служить многообразные приспособления цветка к перекрестному опылению ветром и насекомыми, приспособления плодов и семян к рассеиванию, приспособления листьев к уменьшению испарения.
Обычно мелкие систематические группы – виды, роды, семейства – в процессе эволюции возникают путем идиоадаптаций.
С появлением человека причины биологического прогресса и биологического регресса часто связаны с изменениями, которые он вносит в ландшафты Земли, нарушая связи живых существ со средой, сложившиеся в процессе эволюции. Деятельность человека является мощным фактором биологического прогресса одних видов, нередко вредных для него, и биологического регресса других, нужных и полезных ему. Вспомните появление многих видов насекомых, устойчивых к ядохимикатам, болезнетворных микробов, устойчивых против лекарств, бурное развитие сине–зелёных водорослей в сточных водах. Виной всему этому деятельность человека. При посевах и посадках он вторгается в живую природу, уничтожает на больших массивах великое многообразие диких популяций, заменяя их немногими искусственными. Усиленное истребление человеком многих видов ведет их к биологическому регрессу. Биологический регресс всегда грозит вымиранием. Вот почему мероприятиями по охране природы важно не только сдерживать , но и предупреждать. В хозяйственной деятельности и в медицине человеку необходимо учитывать биологический прогресс и биологический регресс.
Доказательства эволюции: эмбриологические, палеонтологические, сравнительно–анатомические,
Эмбриологические доказательства
.Убедительные доказательства степени родства между организмами представляет эмбриология, изучающая зародышевое развитие организмов. Еще Ч. Дарвин отметил наличие взаимосвязей между индивидуальным развитием организмов (онтогенезом)
иих эволюционным развитием (филогенезом)
.
Сходство зародышей.
Подавляющее большинство организмов развиваются из оплодотворенного яйца. Удивительное сходство зародышей рыбы, ящерицы, кролика, человека касается формы тела, наличия хвоста, зачатков конечностей, жаберных карманов по бокам глотки. Во многом сходна на этих стадиях внутренняя организация зародышей. У всех сначала имеется хорда, а затем позвоночник из хрящевых позвонков, кровеносная система с одним кругом кровообращения (как у рыб), одинаковое строение почек и др. Закон зародышевого сходства, открытый К.Бэром: зародыши позвоночных животных сходны, особенно на более ранних стадиях развития
.
По мере развития сходство между зародышами ослабевает и начинают всё более четко проявляться черты тех классов, к которым они принадлежат. У ящерицы, кролика и человека зарастают жаберные карманы; у зародыша человека особенно сильно развивается головной отдел, включающий мозг, формируются пятипалые конечности, а у зародыша рыбы – плавники и т. п. По ходу эмбрионального развития последовательно происходит расхождение признаков зародышей, приобретающих черты , характеризующие класс, отряд, род и, наконец, вид, к которому они принадлежат.

Биогенетический закон. Во второй половине XIXв. немецкие ученые Ф. Мюллер и Э. Геккель установили закон соотношения онтогенеза, который получил название биогенетического закона. Каждая особь в индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез), или, короче, онтогенез есть краткое повторение филогенеза .
Пример. У всех без исключения позвоночных животных в онтогенезе закладывается хорда – признак, который был свойствен их отдаленным предкам. У головастиков бесхвостых земноводных (лягушки, жабы) развивается хвост. Это – повторение признаков их хвостатых предков. Личинки многих насекомых имеют червеобразную форму (гусеницы бабочек, личинки мух и т. д.). В этом следует усматривать повторение черт строения их червеобразных предков.
Из споры мха развивается сначала ветвящаяся нить, похожая на нитчатую водоросль. Это говорит о родстве наземных растений с водорослями.
Биогенетический закон имеет большое значение для выяснения родственных связей между организмами.
Сопоставление ископаемых остатков из земных пластов разных геологических эпох убедительно свидетельствует об изменении органического мира во времени. В самых древних пластах заключены остатки типов беспозвоночных, а в более поздних пластах – остатки типа хордовых. Позже на Земле появились позвоночные. В более молодых геологических пластах содержатся остатки животных и растений, относящихся к видам, похожим на современные.
Данные палеонтологии дают большой материал о преемственных связях между различными систематическими группами. В одних случаях удалось установить переходные формы, в других – филогенетические ряды, т. е. ряды видов, последовательно сменяющих один другой.
Ископаемые переходные формы. На берегах Северной Двины была найдена группа зверозубых рептилий. Они совмещают признаки пресмыкающихся и млекопитающих. Такие организмы относят к переходным формам. Зверозубые рептилии имеют сходство с млекопитающими в строении черепа, позвоночника и конечностей, а также в делении зубов на клыки, резцы и коренные.


Большой интерес с эволюционной точки зрения представляет находка археоптерикса. Это животное величиной с голубя имело признаки птицы (сходство задних конечностей с цевкой, наличие перьев и общий вид), но сохраняло еще черты пресмыкающихся (длинный ряд хвостовых позвонков, брюшные ребра и наличие зубов). Археоптерикс не мог быть хорошим летуном, так как у него слабо развиты грудная кость (без киля), мышцы крыльев и грудные Позвоночник и ребра не являлись жесткой костной системой, устойчивой при полете , как у современных птиц.
Рыбы → кистепёрые рыбы → земноводные.
Споровые папоротники → семенные папоротники → голосеменные.
Земноводные → стегоцефалы → пресмыкающиеся.
Одноклеточные → вольвокс → многоклеточные.
Одноклеточные растения → эвглена зелёная → одноклеточные животные.
Филогенетические ряды. Последовательные ряды ископаемых форм, последовательно сменяющие друг друга – филогенетические ряды . Примером может служить эволюция лошади.
Наиболее древний ее предок ростом с лисицу, с четырехпалыми передними конечностями, трехпалыми задними и бугорчатыми зубами травоядного типа. Жил в местностях с теплым и влажным климатом, среди трав и кустарников, передвигался скачками. К концу неогена растительность стала более сухой и грубой; в открытых степных пространствах спасение от врагов можно было найти в быстром беге, других средств защиты у этих животных не было.

Борьба за существование и естественный отбор проходили в направлении удлинения ног и сокращения поверхности опоры – уменьшения количества пальцев, достигающих почвы, упрочения позвоночника, что способствовало быстрому бегу. Изменение характера пищи повлияло на образование складчатых зубов. В результате произошла мощная перестройка организма этих животных.
По мере перехода от более древних земных слоев к новым наблюдается постепенное повышение организации животных и растений, постепенное приближение фаун и флор к современным.
Сравнительно–анатомические
доказательства
. К ним относятся:
клеточное строение организма;
общий план строения позвоночных животных;
рудименты (органы, которые выполняли определённые функции у предков, а у потомков находятся в стадии исчезновения или выполняют новые функции – аппендикс, третье веко, копчик у человека);
атавизмы (случаи возврата к признакам предков – у человека многососковость, сильно развитый волосяной покров на теле);
гомологичные органы (имеют общее происхождение , сходное строение и расположение, но выполняют разные функции – лепестки у кувшинок гомологичны тычинкам, гомологичны конечности насекомых и позвоночных)
аналогичные органы (не имеют общего плана строения и общего происхождения, но выполняют сходные функции – крыло бабочки и птицы, жабры рыбы и речного рака);
дивергенция – расхождение признаков в пределах популяции, вида (заяц–беляк и заяц–русак, клевер луговой и клевер ползучий);
конвергенция – сближение признаков в пределах систематических групп (форма тела кита и рыбы, корень и корневище).
Биогеографические доказательства . Изучает сходство и различие флоры и фауны различных материков, островных и материковых видов т.д.
Доказательства эволюции


Фронтальный опрос:
1. Дайте определение понятиям «макроэволюция», «биологический прогресс», «биологический регресс», «рудименты», «атавизмы», «аналогичные органы», «гомологичные органы», «дивергенция», «конвергенция».
2. Назовите и охарактеризуйте пути достижения биологического прогресса.
3. Перечислите и охарактеризуйте доказательства эволюции.
А). В чем выражаются сходство и различие зародышей позвоночных.
Б). Какие доказательства эволюции представляет палеонтология?
в). Приведите примеры современных и ископаемых переходных форм. Какое значение имеют ископаемые формы для изучения органического мира?
Г). Какое значение имеют исследования филогенетических рядов?
4. Приведите примеры биологического прогресса и биологического регресса.
II. Представления о происхождении жизни на Земле.
Геологические эры.
Гипотезы о происхождении жизни на Земле
При развитии взглядов о происхождении жизни возникли теории биогенеза («живое возникает из живого»), абиогенеза («живое возникает из неживого») и допускали самозарождение жизни. В XVIIв. Франческо Реди провёл опыт доказывающий, что «живое возникает из живого. Поместил гнилое мясо в закрытый сосуд, на нём не произошло развития личинок мух. Противники считали, что это не произошло вследствие того, что не проходил воздух. Тогда Реди взял несколько открытых сосудов, а один прикрыл марлей. Через некоторое время в открытых сосудах мясо кишело личинками мух, тогда как в сосудах, прикрытых марлей, в гнилом мясе никаких личинок не было. Но так и не доказал, т.к. сторонники виталистического учения считали, что в каждом организме есть «жизненная сила», после того как ее «вдохнуть» в неживое , возникает живое.
Эксперимент Луи Пастера
. Он кипятил в колбе различные питательные среды, в которых могли развиваться микроорганизмы. При длительном кипячении в колбе погибали не только микроорганизмы, но и их споры. Помня об утверждении виталистов, что мифическая «жизненная сила» не может проникнуть в запаянную колбу, Пастер присоединил к ней S–образную трубку со свободным концом.. Споры микроорганизмов оседали на поверхности тонкой изогнутой трубки и не могли проникнуть в питательную среду. Хорошо прокипячённая питательная среда оставалась стерильной, в ней не наблюдалось самозарождения микроорганизмов, хотя доступ воздуха был обеспечен. Стоило, однако, колбу повернуть так, чтобы содержащийся в ней бульон обмыл S–образное колено горлышка и стек обратно в колбу, в ней вскоре начиналось гниение. Это происходило потому, что в бульон попадали микробы, находившиеся в S–образной части горлышка. Таким образом
, невозможность самопроизвольного зарождения микроорганизмов была убедительно доказана.

Современные представления о происхождении жизни.
Современные теории происхождения жизни исходят из положения о том, что происхождение и эволюция живого определяется химическим составом древней Земли. Экспериментальное подтверждение получила гипотеза А.И.Опарина. Основные положения гипотезы (1922г):
Условия возникновения жизни на Земле (разогревание Земли, ускорение химических реакций, образование первичной атмосферы, своеобразие её газового состава, охлаждение планеты, появление первобытного океана);
Абиогенный синтез простейших органических веществ из неорганических, используемые при этом источники энергии. Возможные источники энергии: молнии, УФИ, космические лучи, радиоактивные частицы , ударные волны от метеоритов, тепло от вулканов, гейзеров, горячих источников;
Образование белков, жиров, углеводов, нуклеиновых кислот;
Образование коацерватов как обособление в растворе более высокомолекулярных веществ в виде высококонцентрированного раствора;
Взаимодействие коацерватов с окружающей средой, сходство с живыми организмами: рост, питание, дыхание, обмен веществ, размножение;
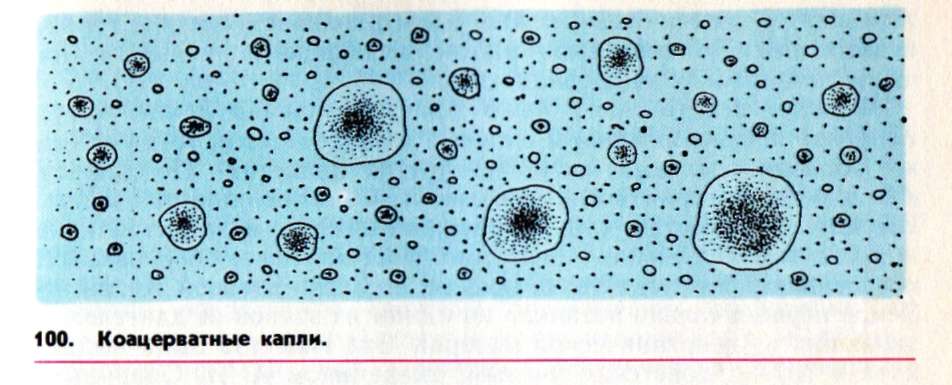
Возникновение полинуклеотидов, способных воспроизводить себе подобных.
1. С.Аррениус и В.И.Вернадсткий – жизнь существует вечно и переносится космическими лучами с одной планеты на другую, пока не встретит подходящие условия для своего развития;
2. Либих – теория «панспермии» - с планеты на планету с метеоритами заносятся простейшие организмы и споры;
3. Крик и Орчел – Земля «засеяна» существами из других планетных систем с помощью ракет с контейнерами простейших организмов;
4. И.С.Шкловский – наша цивилизация является единственной в нашей Галактике или даже во всей наблюдаемой части Вселенной.
Гипотезы, которая могла бы стать «руководящей» и превратиться во всеобъемлющую теорию , пока ещё нет.
Геологические эры.
Наукой установлено, что жизнь возникла в океане около 3,5 млрд. лет назад. Все первые этапы развития жизни протекали в водной стихии. Выход организмов на сушу осуществился примерно 2–2,5 млрд. лет назад. Этому способствовал важный ароморфоз у растений – образование органов и тканей, что имело решающее значение в эволюции растительного мира.
В условиях наземного существования растений дальнейшее развитие растительного мира было связано с другим крупным ароморфозом – переходом от размножения спорами к размножению семенами. Голосеменные растения достигли значительного развития в конце палеозойской эры в пермском периоде. Господство голосеменных в наземной флоре относится к первой половине мезозойской эры, особенно к юрскому периоду. На смену голосеменным в результате новых ароморфозов приходят покрытосеменные растения. Покрытосеменные становятся господствующими на Земле, приспособленными к самым различным условиям существования.
Многочисленные ароморфозы отмечены и в эволюции животных. Многие из них были связаны с переходом к наземному существованию. Крупным ароморфозом при переходе на сушу явилось развитие внутреннего оплодотворения и ряд приспособлений к развитию зародыша в яйце на суше. Птицы и млекопитающие заняли господствующее положение среди наземных позвоночных. Постоянная температура тела позволила им выжить в условиях оледенения и проникнуть далеко в холодные страны. Успешному развитию обеих групп способствовали ароморфозы и идиоадаптации, которые позволили млекопитающим освоить наземную, а птицам – воздушную среду.
Особенно важными ароморфозами в эволюции позвоночных явилось преобразование головного мозга, прогрессивное развитие коры больших полушарий. Все это резко повысило уровень нервной деятельности, усложнило систему условных рефлексов и форм поведения животных в природе. От животных предков ароморфная эволюция привела к возникновению человека.
В антропогене животный мир принимает современный облик.

Геохронологическая таблица


Геологические эры и периоды указаны в таблице соответственно расположению земных пластов: позднейшие – наверху, древнейшие – внизу. Поэтому чтение таблицы начинайте снизу – с архейской эры – и последовательно переходите к более поздним эрам и периодам.
Фронтальный опрос:
1. Гипотезы о происхождении жизни на Земле. Опыты Ф.Реди и Л.Пастера
2. Современные представления о происхождении жизни.
3. Теория Опарина
4. Геологические эры.
III. Итоговое занятие по разделу:
«Развитие органического мира».
Понятие макроэволюции.
Дайте определение понятию и охарактеризуйте «биологический регресс».
Дайте определение понятию и охарактеризуйте «биологический прогресс».
Главные направления биологического прогресса: ароморфоз, идиоадаптация, дегенерация. Приведите примеры.
Охарактеризуйте и приведите примеры доказательств эволюции: эмбриологических, палеонтологических, сравнительно–анатомических, биохимических, биогеографических.
Гипотезы о происхождении жизни на Земле (Опыты Ф.Реди и Л.Пастера).
Современные представления о происхождении жизни. Теория Опарина.
Геологические эры.
Контрольная работа
I вариант
1. Дайте определение понятиям «биологический прогресс», «микроэволюция», приведите примеры.
2. Теория Опарина.
3. Палеонтологические доказательства эволюции.
4. Ароморфоз. Дегенерация.
II вариант
Дайте определение понятию «биологический регресс», «макроэволюция», приведите примеры.
Опыты Ф.Реди и Л.Пастера.
Эмбриологические доказательства эволюции.
Идиоадаптация. Дегенерация.
"Развитие органического мира"
Море - первичная среда развития жизни
Ранние этапы развития жизни на Земле. Вместе с другими планетами солнечной системы Земля образовалась, как предполагают, 5-7 млрд. лет назад. Многие сотни миллионов лет условия, необходимые для жизни, отсутствовали. Это была звездная эра в истории Земли.
Первая геологическая эра - архейская, продолжительностью 900 млн. лет, - почти не оставила следов органической жизни.
Это объясняется тем, что осадочные слои архейского возраста были сильно видоизменены под действием высоких температур и давления, а также тем, что твердые части тела у первичных организмов были развиты слабо.
Однако наличие пород органического происхождения - известняка, мрамора и углистых веществ - указывает на то, что в архейскую эру уже существовали живые организмы: бактерии, водоросли:
Жизнь сосредоточивалась в воде, которую водоросли уже насытили кислородом, что и создавало условия для появления животных.
Таким образом, к началу протерозойской эры (длившейся 2000 млн. лет) произошли три крупных изменения в развитии живых организмов - возникновение:
1) полового процесса,
2) фотосинтеза и
3) многоклеточное.
Половой процесс возник, как предполагают, в форме слияния двух одинаковых клеток, например двух особей одноклеточных водорослей. Позднее половое размножение, по-видимому, происходило при помощи специальных половых клеток.
При половом процессе мужская и женская половые клетки сливаются, образуя зиготу. Из нее развивается организм, совмещающий наследственность как отца, так и матери, это усиливает наследственную изменчивость в потомстве, а следовательно, расширяет действие естественного отбора. Новый способ размножения, как полезный в сохранении видов, был закреплен естественным отбором, и теперь он преобладает в животном и растительном мире.
Возникновение фотосинтеза положило начало разделению единого ствола жизни на два - растения и животные. На самых ранних этапах эволюции произошла дивергенция организмов по способу питания.
Первичные организмы могли существовать за счет разложения органических веществ, находившихся в изобилии в древнейших водных бассейнах. Некоторые первичные организмы получали необходимую для жизни энергию за счет той, которая выделялась при химических реакциях.
И в наше время существуют бактерии, которые используют энергию, освобождаемую при окислении, например, азота, серы, железа.
Организмы, начавшие синтезировать пигмент хлорофилл, получили возможность использовать запасы углерода в форме углекислого газа и перерабатывать их в органические вещества на солнечном свету. В атмосфере появился свободный кислород.
Изменение воздушной среды и наличие пищи - зеленых растений - создали предпосылки для развития животных.
Родоначальниками простейших растений и животных считается группа древнейших одноклеточных организмов - жгутиковых. Бактерии и сине-зеленые водоросли не дали начала новым типам и остались до нашего времени обособленными группами.
Предками многоклеточных организмов, как предполагают многие ученые, были колониальные формы одноклеточных жгутиковых.
И в настоящее время существуют колониальные формы жгутиковых. Многоклеточное строение повлекло дальнейшее усложнение в организации живых существ - дифференциацию тканей, органов и их функций.
Остатки животного мира протерозойской эры крайне редки и малочисленны, но по ним можно судить о том, какой бесконечно длинный путь развития был уже пройден органическим миром. Известны остатки всех типов беспозвоночных животных, включая высокоразвитые типы: иглокожих и членистоногих. Предполагают, что в конце протерозойской эры появились первичные хордовые - подтип бесчерепных, единственным представителем которых в современной фауне является ланцетник.
Важнейшим этапом было возникновение животных с двусторонней симметрией тела, которая привела к дифференцировке его на передний и задний концы, а также на брюшную и спинную стороны.
Передний конец является местом, где развиваются органы чувств, нервные узлы, а в дальнейшем - головной мозг; спинная сторона выполняет защитную функцию; брюшная обеспечивает захват пищи. Подавляющее большинство многоклеточных животных двусторонне симметричны.
Растения и животные в протерозойскую эру еще не заселили сушу, но бактерии и одноклеточные водоросли могли существовать во влажных местах, принимая участие в процессах почвообразования.
Палеозойская эра. Начиная с кембрийского периода, эта эра отличается большим разнообразием форм. Кроме бактерий и одноклеточных водорослей, получили распространение крупные многоклеточные водоросли
В эволюционном отношении многоклеточные водоросли представляли большой шаг вперед, так как обладали более совершенными способами питания и размножения по сравнению с одноклеточными водорослями.
Расчленение тела у многоклеточных водорослей путем ветвления и образования различных выростов, напоминавших по виду листья, стебли и корни, увеличивало поверхность соприкосновения с внешней средой, откуда они получали питательные вещества и воду.
О фаунах кембрийского и силурийского периодов свидетельствуют ископаемые остатки, относящиеся ко всем современным типам: простейшим, кишечнополостным, губкам, червям (три типа), иглокожим, моллюскам, членистоногим, хордовым.
В морях силурийского периода процветали плеченогие. Это сидячие небольшие животные (до 10 см в длину) с двустворчатой раковиной; в современной фауне их осталось не более 200 видов. В илистом грунте ползали или плавали у самого дна представители членистоногих - трилобиты, которые долгое время оставались наиболее высокоорганизованными животными.
Самые крупные трилобиты достигали 75 см в длину. Обнаружены остатки бесчелюстных позвоночных - щитковых. Это небольшие животные с внутренним хрящевым скелетом и твердым внешним панцирем. Современные миноги и миксины - отдаленные потомки щитковых.
У представителей этих фаун было настолько сложное строение, что возникает предположение о существовании их предков еще в протерозойскую эру, но до нас не сохранившихся.
В процессе дальнейшей эволюции происходила главным образом дивергенция типов животного мира и замена первоначальных низкоорганизованных, примитивных форм более высокоорганизованными.
Развитие наземных организмов в палеозойскую эру
Выход растений на сушу. В конце силурийского периода в связи с бурными горообразовательными процессами большая территория суши освободилась из-под воды. Многие виды водорослей, оказавшиеся в условиях суши, погибли.
Но какая-то часть зеленых многоклеточных водорослей приобрела изменения, имевшие широкое приспособительное значение в новой среде обитания. Растениями, вступившими на новый путь развития, были псилофиты.
По сравнению с зелеными многоклеточными водорослями псилофиты претерпели крупнейшие изменения. У них возникли ткани, выполняющие различные функции: защитная (эпидермис с устьицами), механическая, поддерживающая тело в пространстве.
Ветвление стебля продолжалось под землей, образуя подобие корней.
Возможность заселения суши псилофитами была уже подготовлена жизнедеятельностью бактерий, одноклеточных водорослей, создавших первую почву.
С середины девонского периода происходит постепенное убывание псилофитов и их полное исчезновение к концу этого периода. Они дали начало группам растений, стоящим выше по степени организации: плауновым, хвощовым и папоротниковым.
Каменноугольные леса. В развитии наземной растительности произошел крупный эволюционный подъем около 350 млн. лет назад, в начале каменноугольного периода с его теплым влажным климатом и воздухом, богатым углекислым газом вследствие сильной вулканической деятельности.
Впервые на Земле появились огромные леса из папоротникообразных - гигантских папоротников, древовидных хвощовых и плауновых - высотой 15-30 м. В подлеске произрастали травянистые формы папоротникообразных; стволы обвивали лианы - папоротники.
По сравнению с псилофитами папоротникообразные обладали многими преимуществами: хорошим развитием корневой и проводящей систем, а также - листьев. Но в размножении папоротникообразные все еще были связаны с водой, необходимой для передвижения половых клеток при оплодотворении.
Пышная растительность сильно изменила состав атмосферы, обогатив ее кислородом, что имело важное значение для развития наземных животных.
Растительность каменноугольного периода способствовала образованию плодородных почв, обогащая их перегноем. А это благоприятствовало дальнейшему развитию наземной растительности.
Леса каменноугольного периода образовали месторождения каменного угля, имеющие исключительное значение для нас.
Древнейшие семенные растения. В каменноугольный период в лесах произрастали семенные папоротники - древнейшие голосеменные, у которых вместо спорангиев со спорами развивались семена. Семенные папоротники ясно указывают на происхождение голосеменных растений из споровых.
Возникновение первых семенных растений было огромным прогрессивным фактом, определившим всю дальнейшую эволюцию растений.
Особенности размножения семенных растений создавали им преимущественное положение по сравнению с папоротникообразными. У семенных растений оплодотворение происходит без участия воды, зародыш находится в семени и поэтому защищен от неблагоприятных влияний, семя содержит большой запас питательных веществ, необходимых для развития зародыша.
Папоротникообразные же нуждаются в наличии воды для оплодотворения; спора у них представляет собой всего одну клетку с очень ограниченным запасом питательных веществ; для развития заростка необходима влажная почва.
С конца каменноугольного периода в связи с усиленным горообразованием, охватившим в следующем (пермском) периоде весь земной шар, влажный климат почти повсеместно сменился сухим. В новых условиях древовидные папоротникообразные стали быстро вымирать; лишь в сырых и тенистых местах удержались более мелкие формы.
Вымерли и семенные папоротники. Но более высокоорганизованные голосеменные растения оказались жизнестойкими, так как размножение семенами позволяло расселяться на более сухих местах. Богатое развитие и широкое распространение голосеменных продолжалось почти до конца мезозойской эры.
Выход позвоночных на сушу. Появление и развитие наземных растений создало важнейшие предпосылки для выхода животных на сушу и дальнейшего развития их в новых условиях; изменение химического состава атмосферы и запасов пищи.
Чтобы понять, как совершился выход животных на сушу, следует остановиться на исключительно важных в эволюционном отношении изменениях, происшедших у рыб в девонском периоде. Первое крупное преобразование связано с появлением челюстных панцирных рыб. Считают, что их предками была одна из групп щитковых.
Челюстные панцирные рыбы обладали внутренним хрящевым скелетом и подвижными костными челюстями, парными плавниками и костным панцирем.
Возникновение челюстей было важным этапом по пути повышения общей организации позвоночных.
Животные, обладавшие костными челюстями, могли активнее охотиться, энергичнее справляться с добычей, в связи с чем в процессе естественного отбора совершенствовались нервная система и органы чувств.
В девонском периоде появилась еще новая группа рыб - кистеперые. Они имели два способа дыхания: жаберное и легочное при помощи одного или двух пузырей, открывающихся на брюшной стороне пищевода.
Плавники кистеперых не были специализированы, как, например, у челюстных панцирных рыб, в выполнении одной функции, т.е. плавания. Плавники кистеперых выполняли несколько функций: они служили не только при плавании, но и порой при ползании. Скелет плавника кистеперой рыбы гомологичен пятипалой конечности позвоночных.
Кистеперые обитали в пресных водоемах. В мезозойскую эру они перешли в моря. В 1938 г. в Индийском океане, у берегов Африки, было поймано несколько экземпляров рыбы, названной латимерией. Это дошедшие до нас представители кистеперых рыб, считавшихся полностью вымершими.
Самыми примитивными наземными позвоночными считают древних земноводных. Установлено, что они произошли от одной из групп кистеперых рыб. Процесс возникновения земно-74 водных можно представить себе таким образом. Рыбы, принадлежавшие к одной из ветвей кистеперых, были крупнее других и обладали более мощными плавниками.
Они жили в неглубоких водоемах, периодически высыхавших, что заставляло животных переползать в оставшиеся лужи. На основе наследственной изменчивости и в процессе естественного отбора плавники превратились в конечности, пригодные для хождения.
Предпосылками для выхода на сушу кистеперых предков земноводных были строение конечностей и легочное дыхание.
Древнейшие земноводные - стегоцефалы (плоскоголовые) обитали в болотистых местностях. Размножение, развитие яиц и личинок, дышавших жабрами, происходило, как и у всех современных земноводных, в воде.
Поэтому первые наземные животные не могли жить в местах, удаленных от водоемов. По сравнению с современными земноводными их далекие предки отличались разнообразием размеров (от нескольких сантиметров до 4 м в длину), наличием костных щитков на голове и мелких щитков на брюхе.
Стегоцефалы соединяли признаки рыб, земноводных и пресмыкающихся. Это переходная форма.
Теплый и влажный климат, огромные леса и топи каменноугольного периода создавали исключительно благоприятные условия для расцвета групп земноводных, приспособленных к различным местообитаниям и находящихся на одном уровне общей организации.
В каменноугольный период на суше шло интенсивное и разностороннее развитие беспозвоночных: насекомых, пауков, скорпионов.
В морях плавало множество рыб, особенно акул. Кораллы продолжали возводить свои известковые постройки; из раковин корненожек шло интенсивное образование горных пород. Дно усеивали плеченогие, моллюски, иглокожие; заметно убавилось трилобитов.
Ранее существовавшие типы и классы дивергировали на новые группы; вымирали одни ветви, происходил подъем и расцвет других.
Естественный отбор "прилаживал" живые существа к разным местообитаниям, хотя и в пределах одной среды; это ослабляло действие конкуренции и способствовало сохранению видов.
В конце каменноугольного периода, особенно в начале пермского, создались засушливые условия, крайне неблагоприятные для земноводных. Прежде всего, их размножение зависело от наличия воды, без которой не мог развиваться зародыш.
Легочное дыхание, кровеносная система были несовершенными, а голая кожа не могла защищать тело от высыхания. Значительная часть крупных земноводных вымерла.
Те же, которые могли укрыться в оставшихся болотах и топях, дали начало земноводным значительно меньших размеров.
Способной к дальнейшему завоеванию суши оказалась какая-то группа земноводных, которая претерпела очень большие изменения, оказавшиеся полезными в новых условиях.
У этих животных изменился способ размножения: возникло внутреннее оплодотворение; яйцо имело большой запас питательных веществ и оболочку, предохраняющую зародыш от высыхания.
Развитие зародыша происходило на суше. Эти важные преобразования положили конец зависимости размножения и развития животных от воды.
У взрослых животных развился роговой покров, защищавший их от высыхания. Легкие и кровеносная система также оказались более совершенными у этих животных при переходе их полностью к жизни на суше. Таким образом, возник новый класс позвоночных - пресмыкающиеся.
Появление древнейших пресмыкающихся было новым важным этапом в развитии животного мира: из низкоорганизованных земноводных возникли более высокоорганизованные пресмыкающиеся. И когда в конце палеозойской эры климат на Земле стал жарким и засушливым, пресмыкающиеся оказались вне конкуренции в новой среде обитания.
Главным образом это были травоядные животные, но некоторые пресмыкающиеся перешли к хищному образу жизни. Особенно интересна группа зверозубых рептилий, остатки которых были найдены в СССР на берегах Северной Двины.
У них есть сходство с млекопитающими в строении черепа, позвоночника и конечностей, а также делении зубов на клыки, резцы и коренные. Предполагают, что от потомков зверозубых рептилий взяли начало первые млекопитающие. Зверозубые ящеры - переходная форма.
Итак, в палеозойскую эру происходило дальнейшее развитие беспозвоночных в воде; возникли бесчелюстные и челюстные панцирные рыбы.
Растения и животные вышли на сушу; появились сосудистые споровые и голосеменные растения, кистеперые рыбы, земноводные и пресмыкающиеся. В органическом мире происходили глубокие изменения; в разные геологические эры и периоды существовали различные флоры и фауны, преемственно сменявшие друг друга.
Как вы видите, ознакомление с наиболее важными этапами развития органического мира в первые три эры - архейскую, протерозойскую и палеозойскую - дает богатый фактический материал, на основании которого можно судить о направлениях эволюции.
Рассмотрим этот вопрос, прежде чем перейти к изучению дальнейших изменений в живой природе в течение мезозойской и кайнозойской эр.
Список литературы
1. Азимов А. Краткая история биологии. М., 1997.
2. Кемп П., Армс К. Введение в биологию. М., 2000.
3. Либберт Э. Общая биология. М., 1978.
4. Льоцци М. История физики. М., 2001.
5. Найдыш В.М. Концепции современного естествознания. Учебное пособие. М., 1999.
400. Некоторые виды вымирали, сменяясь новыми. И всегда менее специализированные группы в силу большей пластичности своей организации оказывались способными к дальнейшим ароморфозам. Органический мир в кайнозойскую эру Господство покрытосеменных. Кайнозойская эра длится около 60-70 млн. лет. Первый период ее - палеоген, второй - неоген и третий - антропоген, который длится по настоящее...
Рудиментов возможно объяснить только тем, что эти органы у далеких предков были нормально развиты и функционировали, но в процессе эволюции потеряли свое биологическое значение и сохранились в виде остатков. Рудименты служат важными доказательствами исторического развития органического мира. Ни один из противников Дарвина не пытался ссылаться на рудименты в своих возражениях; слишком нелепо было...
Эволюционное развитие органического мираРаздел: «Развитие органического мира» (6 часов)
Эволюционное развитие органического мира.
I. Главные пути и направления эволюции органического мира. Макроэволюция, её доказательства.
Доказательства эволюции: эмбриологические, палеонтологические, сравнительно-анатомические, биохимические, биогеографические.
^ Понятие макроэволюции.
Процесс образования из видов новых родов, из родов - новых семейств и т. п. называют макроэволюцией . Она происходит в исторически грандиозные промежутки времени и недоступна непосредственному изучению. Макроэволюция - надвидовая эволюция, в отличие от микроэволюции, происходящей внутри вида, внутри его популяций. Однако принципиальных отличий между этими процессами нет, так как в основе макроэволюционных процессов лежат микроэволюционные. В макроэволюции действуют те же процессы - борьба за существование, естественный отбор и связанное с ними вымирание. Макроэволюция носит дивергентный характер, так же как и микроэволюция.
^ Понятие биологического регресса.
Биологический регресс - направление эволюции, которое приводит к сокращению численности вида, сужению его ареала, уменьшению числа популяций вида и, возможно, в конечном счете, к его гибели. Причины биологического регресса: деятельность человека, приводящая к непосредственному или опосредованному уничтожению видов; эволюционные ограничения - возможности эволюционных изменений вида не безграничны, т.к. возникновение ряда мутаций невозможно из-за отсутствия генетических предпосылок. Например, у дрозофил отсутствуют гены, определяющие появление особей с синими или зелёными глазами.
^ 3. Понятие биологического прогресса.
Биологический прогресс - направление эволюции, которое приводит к увеличению численности вида, расширению его ареала, увеличению числа популяций вида и образованию новых популяций вида. Биологический прогресс достигается тремя путями: ароморфозом, идиоадаптацией и дегенерацией.
^ Главные направления биологического прогресса:
ароморфоз, идиоадаптация, дегенерация.
Ароморфоз (арогенез, морфофизиологический прогресс) - эволюционные изменения, которые ведут к общему подъему организации, повышают интенсивность жизнедеятельности, но не являются узкими приспособлениями к резко ограниченным условиям существования. Ароморфоз дает значительные преимущества в борьбе за существование и делает возможным переход в новую среду обитания, способствует повышению выживаемости и снижению смертности в популяции. Так, усложнение строения древних земноводных по сравнению с рыбами способствовало вступлению земноводных на путь биологического прогресса. Например, у млекопитающих сердце разделено на две половины - правую и левую с развитыми двумя кругами кровообращения. Дифференцировались органы пищеварения.
Идиоадаптация (аллогенез) - мелкие эволюционные изменения, которые способствуют приспособлению к определенным условиям среды обитания (частные приспособления). Идиоадаптация не сопровождается изменениями основных черт организации, общим подъемом её уровня и интенсивности жизнедеятельности организмов. Например, в засушливые и жаркие периоды у некоторых земноводных на основе наследственных изменений в процессе борьбы за существование и естественного отбора выработались частные приспособления к определенным и ограниченным местообитаниям - в болотах и топях. Никакого повышения уровня организации у земноводных при этом не произошло. Современные амфибии - лягушки, тритоны - хорошо приспособлены к условиям существования в мелких водоемах и сильно увлажненных местах суши. Приспособления придонных рыб - камбаловых, скатов к жизни на дне (уплощение тела, окраска под цвет грунта) представляют собой типичные примеры идиоадаптации.
Защитная окраска дает хороший пример приспособления животных к определенным условиям существования, не связанного с повышением организации.
Примерами идиоадаптации у растений могут служить многообразные приспособления цветка к перекрестному опылению ветром и насекомыми, приспособления плодов и семян к рассеиванию, приспособления листьев к уменьшению испарения.
Обычно мелкие систематические группы - виды, роды, семейства - в процессе эволюции возникают путем идиоадаптаций.
^ Биологический прогресс и биологический регресс при участии деятельности человека. Из многочисленных ветвей древнейших земноводных остались только те, которые повели к образованию современных классов земноводных и пресмыкающихся. Исчезли древние папоротникообразные и многие другие группы растений и животных.
С появлением человека причины биологического прогресса и биологического регресса часто связаны с изменениями, которые он вносит в ландшафты Земли, нарушая связи живых существ со средой, сложившиеся в процессе эволюции.
^ Доказательства эволюции: эмбриологические, палеонтологические, сравнительно-анатомические,
биохимические, биогеографические.
Эмбриологические доказательства.Убедительные доказательства степени родства между организмами представляет эмбриология, изучающая зародышевое развитие организмов. Еще Ч. Дарвин отметил наличие взаимосвязей между индивидуальным развитием организмов (онтогенезом) иих эволюционным развитием (филогенезом) .
Сходство зародышей. Подавляющее большинство организмов развиваются из оплодотворенного яйца. Удивительное сходство зародышей рыбы, ящерицы, кролика, человека касается формы тела, наличия хвоста, зачатков конечностей, жаберных карманов по бокам глотки. Во многом сходна на этих стадиях внутренняя организация зародышей. У всех сначала имеется хорда, а затем позвоночник из хрящевых позвонков, кровеносная система с одним кругом кровообращения (как у рыб), одинаковое строение почек и др. Закон зародышевого сходства, открытый К.Бэром: зародыши позвоночных животных сходны, особенно на более ранних стадиях развития.
По мере развития сходство между зародышами ослабевает и начинают всё более четко проявляться черты тех классов, к которым они принадлежат. У ящерицы, кролика и человека зарастают жаберные карманы; у зародыша человека особенно сильно развивается головной отдел, включающий мозг, формируются пятипалые конечности, а у зародыша рыбы - плавники и т. п. По ходу эмбрионального развития последовательно происходит расхождение признаков зародышей, приобретающих черты, характеризующие класс, отряд, род и, наконец, вид, к которому они принадлежат.
^ Биогенетический закон. Во второй половине XIXв. немецкие ученые Ф. Мюллер и Э. Геккель установили закон соотношения онтогенеза, который получил название биогенетического закона. Каждая особь в индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез), или, короче, онтогенез есть краткое повторение филогенеза.
Пример. У всех без исключения позвоночных животных в онтогенезе закладывается хорда - признак, который был свойствен их отдаленным предкам. У головастиков бесхвостых земноводных (лягушки, жабы) развивается хвост. Это - повторение признаков их хвостатых предков. Личинки многих насекомых имеют червеобразную форму (гусеницы бабочек, личинки мух и т. д.). В этом следует усматривать повторение черт строения их червеобразных предков.
Из споры мха развивается сначала ветвящаяся нить, похожая на нитчатую водоросль. Это говорит о родстве наземных растений с водорослями.
Биогенетический закон имеет большое значение для выяснения родственных связей между организмами.
^ Палеонтологические доказательства.Палеонтология изучает ископаемые остатки вымерших организмов и выявляет их сходство и различие с современными организмами. По ископаемым остаткам палеонтологи восстанавливают внешний вид и строение вымерших организмов, узнают о растительном и животном мире прошлого.
Сопоставление ископаемых остатков из земных пластов разных геологических эпох убедительно свидетельствует об изменении органического мира во времени. В самых древних пластах заключены остатки типов беспозвоночных, а в более поздних пластах - остатки типа хордовых. Позже на Земле появились позвоночные. В более молодых геологических пластах содержатся остатки животных и растений, относящихся к видам, похожим на современные.
Данные палеонтологии дают большой материал о преемственных связях между различными систематическими группами. В одних случаях удалось установить переходные формы, в других - филогенетические ряды, т. е. ряды видов, последовательно сменяющих один другой.
^ Ископаемые переходные формы. На берегах Северной Двины была найдена группа зверозубых рептилий. Они совмещают признаки пресмыкающихся и млекопитающих. Такие организмы относят к переходным формам. Зверозубые рептилии имеют сходство с млекопитающими в строении черепа, позвоночника и конечностей, а также в делении зубов на клыки, резцы и коренные.
Большой интерес с эволюционной точки зрения представляет находка археоптерикса.
Рудименты (органы, которые выполняли определённые функции у предков, а у потомков находятся в стадии исчезновения или выполняют новые функции - аппендикс, третье веко, копчик у человека);
Гомологичные органы (имеют общее происхождение, сходное строение и расположение, но выполняют разные функции - лепестки у кувшинок гомологичны тычинкам, гомологичны конечности насекомых и позвоночных)
Дивергенция - расхождение признаков в пределах популяции, вида (заяц-беляк и заяц-русак, клевер луговой и клевер ползучий);
Конвергенция - сближение признаков в пределах систематических групп (форма тела кита и рыбы, корень и корневище).
^ Биохимические доказательства. Близкородственные виды имеют сходный химический состав. Родство человека и человекообразных обезьян, птиц и крокодилов установлено на основе сходства строения белков и ДНК.
^ Биогеографические доказательства. Изучает сходство и различие флоры и фауны различных материков, островных и материковых видов т.д.
Доказательства эволюции
Фронтальный опрос:
1. Дайте определение понятиям «макроэволюция», «биологический прогресс», «биологический регресс», «рудименты», «атавизмы», «аналогичные органы», «гомологичные органы», «дивергенция», «конвергенция».
2. Назовите и охарактеризуйте пути достижения биологического прогресса.
3. Перечислите и охарактеризуйте доказательства эволюции.
А). В чем выражаются сходство и различие зародышей позвоночных.
Б). Какие доказательства эволюции представляет палеонтология?
В). Приведите примеры современных и ископаемых переходных форм. Какое значение имеют ископаемые формы для изучения органического мира?
Г). Какое значение имеют исследования филогенетических рядов?
4. Приведите примеры биологического прогресса и биологического регресса.
^ II. Представления о происхождении жизни на Земле.
Гипотезы о происхождении жизни на Земле
(Опыты Ф.Реди и Л.Пастера).
При развитии взглядов о происхождении жизни возникли теории биогенеза («живое возникает из живого»), абиогенеза («живое возникает из неживого») и допускали самозарождение жизни. В XVIIв. Франческо Реди провёл опыт доказывающий, что «живое возникает из живого. Поместил гнилое мясо в закрытый сосуд, на нём не произошло развития личинок мух. Противники считали, что это не произошло вследствие того, что не проходил воздух. Тогда Реди взял несколько открытых сосудов, а один прикрыл марлей. Через некоторое время в открытых сосудах мясо кишело личинками мух, тогда как в сосудах, прикрытых марлей, в гнилом мясе никаких личинок не было. Но так и не доказал, т.к. сторонники виталистического учения считали, что в каждом организме есть «жизненная сила», после того как ее «вдохнуть» в неживое, возникает живое.
^ Эксперимент Луи Пастера . Он кипятил в колбе различные питательные среды, в которых могли развиваться микроорганизмы. При длительном кипячении в колбе погибали не только микроорганизмы, но и их споры. Помня об утверждении виталистов, что мифическая «жизненная сила» не может проникнуть в запаянную колбу, Пастер присоединил к ней S-образную трубку со свободным концом.. Споры микроорганизмов оседали на поверхности тонкой изогнутой трубки и не могли проникнуть в питательную среду. Хорошо прокипячённая питательная среда оставалась стерильной, в ней не наблюдалось самозарождения микроорганизмов, хотя доступ воздуха был обеспечен. Стоило, однако, колбу повернуть так, чтобы содержащийся в ней бульон обмыл S-образное колено горлышка и стек обратно в колбу, в ней вскоре начиналось гниение. Это происходило потому, что в бульон попадали микробы, находившиеся в S-образной части горлышка. Таким образом, невозможность самопроизвольного зарождения микроорганизмов была убедительно доказана.
^ Современные представления о происхождении жизни.
Теория Опарина.
Современные теории происхождения жизни исходят из положения о том, что происхождение и эволюция живого определяется химическим составом древней Земли. Экспериментальное подтверждение получила гипотеза А.И.Опарина. Основные положения гипотезы (1922г):
Условия возникновения жизни на Земле (разогревание Земли, ускорение химических реакций, образование первичной атмосферы, своеобразие её газового состава, охлаждение планеты, появление первобытного океана);
Абиогенный синтез простейших органических веществ из неорганических, используемые при этом источники энергии. Возможные источники энергии: молнии, УФИ, космические лучи, радиоактивные частицы, ударные волны от метеоритов, тепло от вулканов, гейзеров, горячих источников;
Взаимодействие коацерватов с окружающей средой, сходство с живыми организмами: рост, питание, дыхание, обмен веществ, размножение;
Возникновение полинуклеотидов, способных воспроизводить себе подобных.
В 1955г. С.Миллер провёл опыт, доказывающий, что первым шагом на Земле был абиогенный синтез органических веществ. Хотя и у этой теории тоже были противники (Фред Хойл). Однако существуют и другие гипотезы зарождения жизни:
1. С.Аррениус и В.И.Вернадсткий - жизнь существует вечно и переносится космическими лучами с одной планеты на другую, пока не встретит подходящие условия для своего развития;
2. Либих - теория «панспермии» - с планеты на планету с метеоритами заносятся простейшие организмы и споры;
3. Крик и Орчел - Земля «засеяна» существами из других планетных систем с помощью ракет с контейнерами простейших организмов;
4. И.С.Шкловский - наша цивилизация является единственной в нашей Галактике или даже во всей наблюдаемой части Вселенной.
Гипотезы, которая могла бы стать «руководящей» и превратиться во всеобъемлющую теорию, пока ещё нет.
^ Геологические эры.
Историю Земли и жизни на ней ученые разбивают на определенные промежутки времени - эры, которые подразделяют на периоды. Результаты исследований, получаемые в различных областях биологической науки, дополняют друг друга. Они позволяют проследить, каким был органический мир в отдаленные эры и периоды, в каких направлениях происходило его развитие, пока он не приобрел современный вид.
Наукой установлено, что жизнь возникла в океане около 3,5 млрд. лет назад. Все первые этапы развития жизни протекали в водной стихии. Выход организмов на сушу осуществился примерно 2-2,5 млрд. лет назад. Этому способствовал важный ароморфоз у растений - образование органов и тканей, что имело решающее значение в эволюции растительного мира.
В условиях наземного существования растений дальнейшее развитие растительного мира было связано с другим крупным ароморфозом - переходом от размножения спорами к размножению семенами. Голосеменные растения достигли значительного развития в конце палеозойской эры в пермском периоде. Господство голосеменных в наземной флоре относится к первой половине мезозойской эры, особенно к юрскому периоду. На смену голосеменным в результате новых ароморфозов приходят покрытосеменные растения. Покрытосеменные становятся господствующими на Земле, приспособленными к самым различным условиям существования.
Многочисленные ароморфозы отмечены и в эволюции животных. Многие из них были связаны с переходом к наземному существованию. Крупным ароморфозом при переходе на сушу явилось развитие внутреннего оплодотворения и ряд приспособлений к развитию зародыша в яйце на суше. Птицы и млекопитающие заняли господствующее положение среди наземных позвоночных. Постоянная температура тела позволила им выжить в условиях оледенения и проникнуть далеко в холодные страны. Успешному развитию обеих групп способствовали ароморфозы и идиоадаптации, которые позволили млекопитающим освоить наземную, а птицам - воздушную среду.
Особенно важными ароморфозами в эволюции позвоночных явилось преобразование головного мозга, прогрессивное развитие коры больших полушарий. Все это резко повысило уровень нервной деятельности, усложнило систему условных рефлексов и форм поведения животных в природе. От животных предков ароморфная эволюция привела к возникновению человека.
В антропогене животный мир принимает современный облик.
Геохронологическая таблица
Геологические эры и периоды указаны в таблице соответственно расположению земных пластов: позднейшие - наверху, древнейшие - внизу. Поэтому чтение таблицы начинайте снизу - с архейской эры - и последовательно переходите к более поздним эрам и периодам.
Фронтальный опрос:
1. Гипотезы о происхождении жизни на Земле. Опыты Ф.Реди и Л.Пастера
2. Современные представления о происхождении жизни.
3. Теория Опарина
4. Геологические эры.
III. Итоговое занятие по разделу:
«Развитие органического мира».
Охарактеризуйте и приведите примеры доказательств эволюции: эмбриологических, палеонтологических, сравнительно-анатомических, биохимических, биогеографических.
Геологические эры.
Контрольная работа
I вариант
1. Дайте определение понятиям «биологический прогресс», «микроэволюция», приведите примеры.
2. Теория Опарина.
3. Палеонтологические доказательства эволюции.
4. Ароморфоз. Дегенерация.
1.6.3.Эволюция органического мира
Эволюция это постепенное длительное развитие органического мира, сопровождающееся его изменением и появлением новых форм организма, при этом развитие идет от простого к сложному . И в далекие геологические эпохи и в настоящее время, органический мир Земли находится в состоянии эволюции. Термин «эволюция» (от лат. развертываю) был предложен в 1762 году швейцарским натуралистом Ш. Боннэ. В биологии вопросы эволюции рассматривает эволюционная теория, которая раньше ограничивалась рамками дарвинизма, а в настоящее время рассматривает как классическое дарвиновское учение, так и современную (синтетическую) теорию эволюции (СТЭ).
Великий английский ученый Чарльз Роберт Дарвин (1809 - 1882), стал творцом первой подлинно научной теории эволюции и внес огромный вклад в создание эволюционного учения, а, следовательно, и развития биологии. Главные работы Ч. Дарвина «Происхождение видов путем естественного отбора…» (1859), «Изменение домашних животных и культурных растений» (1869), «Происхождение человека и половой отбор» (1871).
Рассматривая эволюцию в целом можно увидеть, что результатом ее является все многообразие организмов, живущих на Земле. Поэтому, основываясь на результатах эволюционного процесса можно выделить два вида эволюции - микроэволюцию (эволюция внутри вида) и макроэволюцию (эволюция крупных систематических групп, надвидового ранга).
Микроэволюция - совокупность процессов видообразования, при которых из одного вида возникают новые виды (один или несколько) видов организмов. Примером микроэволюционных процессов является возникновение двух рас ночной бабочки березовая пяденица, разных видов вьюрков на Галапогосских островах, прибрежных видов чаек на побережье Ледовитого океана (от Норвегии до Аляски) и т.д. Выведение породы белой украинской свиньи может служить примером микроэволюции, реализуемой человеком . Образование новых видов из исходного осуществляется за счет дивергенции (см. ниже).
Макроэволюция - совокупность всех эволюционных процессов, результате которых возникло все многообразие органического мира; эти процессы идут не только на уровне вида, но и на уровне рода, семейства, класса и т.д. Результатом макроэволюции является все многообразие современного органического мира, которое возникло как за счет дивергенции, так и за счет конвергенции (схождения признаков). Следовательно, при протекании макроэволюции возможны и дивергенция, и конвергенция (см. ниже).
1.6.3.1. Основные положения эволюционного учения
Эволюционное учение состоит из трех разделов:
- доказательства эволюции
- учение о движущих силах эволюции
- представление о путях эволюционных преобразований
Доказательства эволюции. Выделяют четыре группы доказательств эволюционной теории: цитологические, палеонтологические, сравнительно-анатомические и эмбриологические.
Суть цитологических доказательств состоит в том, что практически все организмы (кроме вирусов) имеют клеточное строение. Клетки и животных, и растений имеют общий план строения и общие по форме и функциям органоиды (цитоплазма, эндоплазматическая сеть, клеточный центр и т.д.). Клетки растений имеют ряд отличий, связанных с различным способом питания и разной приспособленностью к среде обитания по сравнению с животными, однако, существование в природе промежуточного типа одноклеточных организмов - жгутиковых, сочетающих в себе признаки растительных и животных организмов (они как растения способны к фотосинтезу, а как животные - к гетеротрофному способу питания), свидетельствует о единстве происхождения животных и растений. Клетка имеет одинаковый химический и элементарный состав независимо от принадлежности к какому-либо организму, обладая и специфичностью, связанной с особенностью организма.
Эмбриологические доказательства. Первым эмбриологическим доказательством является то, что развитие всех (и животных и растительных) организмов начинается с одной клетки зиготы. Вторым важнейшим доказательством является биогенетический закон (см. тему онтогенез ), согласно которому, онтогенез есть краткое и быстрое повторение филогенеза . Так, отдельные особи вида, независимо от уровня его организации, проходят стадию зиготы, морулы, бластулы, гаструлы, трех зародышевых листков, органогенеза ; более того, и у рыб, и у человека есть личиночная рыбообразная стадия и зародыш человека имеет жабры и жаберные щели (это относится к животным).
Сравнительно-анатомические доказательства эволюции относятся к эволюции животных и основаны на сведениях, полученных сравнительной анатомией. Сравнительная анатомия - наука, изучающая внутреннее строение различных организмов в их сравнении друг с другом (наибольшее значение эта наука имеет для животных и человека). В результате изучения особенностей строения хордовых было обнаружено, что эти организмы имеют двустороннюю (билатеральную) симметрию. Они имеют опорно-двигательную систему, обладающую единым, общим для всех, планом строения (сравните скелет человека и скелет ящерицы или лягушки). Это свидетельствует об общности происхождения человека, пресмыкающихся и земноводных.
Согласно сравнительной анатомии, у различных организмов имеются гомологичные, аналогичные органы, рудименты и атавизмы.
Гомологичными называют органы, имеющие общий план строения, единство происхождения, но они могут иметь различное строение из-за выполнения различных функций. Конечности всех млекопитающих хотя и отличаются друг от друга, но имеют единый план строения и представляют собой пятипалую конечность, примерами являются грудной плавник рыбы, передняя конечность лягушки, крыло птицы и рука человека. Примерами гомологичных органов у растений являются усики гороха, иглы барбариса, колючки кактуса все это видоизмененные листья; корневище ландыша, клубни картофеля, донце репчатого лука подземные побеги, также гомологичны.
Аналогичными называют те органы, которые имеют примерно одинаковое строение (внешняя форма) из-за выполнения близких функций, но обладающих различным планом строения и различным происхождением. К аналогичным органам относится роющая конечность крота и медведки (насекомого, ведущего подземный образ жизни). У организмов, обитающих в воздухе, имеются крылья и другие приспособления для полета, но крылья птицы и летучей мыши измененные конечности, а крылья бабочки выросты стенки тела.
Рудиментами являются остатки тех органов, которые когда-то имели значение, а на данном этапе филогенеза потеряли свое значение. Примерами рудиментов являются аппендикс (слепой отросток кишки), копчиковые позвонки и т.д.
Атавизмы - признаки, ранее присущие и характерные для данного организма, на данном этапе эволюции утратившие свое значение для большинства особей, но проявившиеся у данной конкретной особи в ее онтогенезе. К атавизмам относится хвостатость некоторых людей, полимастия человека (многососковость), чрезмерное развитие волосяного покрова. В отличие от рудиментов, которые присущи всем особям данного вида (например, аппендикс у человека), атавизмы встречаются редко и воспринимаются как уродства, т.е. отклонения в строении.
Палеонтологические исследования позволяют установить историю развития разных форм организмов на Земле, установить родственные (генетические) связи между отдельными организмами, основывается на изучении ископаемых останков.
Учение о движущих силах эволюции. Движущие силы эволюции это те факторы, которые вызывают эволюционный процесс. Эволюционному процессу подвержены э лементарные единицы эволюции.
Элементарными единицами эволюции в теории Ч. Дарвина были виды. Но согласно современным воззрениям, вид не представляет собой наименьшую дискретную, самодостаточную единицу, а является очень сложным образованием, состоящим из отдельных популяций. Поэтому в современной синтетической теории (СТЭ) элементарными единицами эволюции считаются популяции .
Элементарными факторами эволюции , т.е. факторами, приводящими к устойчивому, необратимому, направленному изменению генотипа являются мутации, популяционные волны, изоляция.
Мутации (см. выше) поставляют материал для естественного отбора, приводят к возникновению новых признаков, если признаки благоприятны для организма, они закрепляются в потомстве, накапливаются, что в конечном итоге приводит к появлению новых видов.
Популяционные волны это колебания численности особей в популяции, которые «подставляют» под действие отбора редкие мутации или устраняют обычные варианты.
Изоляция это возникновение различного рода препятствий, делающих невозможным свободное скрещивание, изоляция приводит к закреплению новых свойств и развитию различий.
Также к факторам эволюции относят наследственность, изменчивость, и естественный отбор, который является д вижущей силой эволюции.
Наследственность и изменчивость (см. раздел Генетика) являются важнейшими факторами эволюции. Роль наследственности в эволюции состоит в передаче признаков, в том числе и возникших в онтогенезе, от родителей к потомкам. Изменчивость организмов приводит к появлению особей, имеющих разный уровень отличий друг от друга. Модификационные изменения , не затрагивающие генома, не наследуются. Их роль в эволюции состоит в том, что такие изменения позволяют организму выжить в сложных, порой экстремальных условиях среды. Так, мелкие листья способствуют снижению транспирации (испарения воды), что позволяет растению выжить в условиях недостатка влажности.
Большую роль в процессах эволюции играет мутационная изменчивость , затрагивающая геном гамет. В этом случае возникшие изменения передаются от родителей к потомкам, и новый признак либо закрепляется в потомстве (если он полезен организму), либо организм гибнет, если этот признак ухудшает его приспособленность к среде обитания.
Таким образом, наследственная изменчивость «создает» материал для естественного отбора, а наследственность закрепляет возникшие изменения и приводит к их накапливанию.
Естественный отбор это выживание особей, наиболее приспособленных к конкретным условиям существования и их возможности оставить полноценное, приспособленное к этим условиям существования потомство.
Творческая роль естественного отбора состоит в том, что у организмов возникают признаки, позволяющие наиболее полно приспособиться к данным условиям среды. Например, северный олень, обитающий в полярной тундре выживает только при наличии очень сильных ног, снабженных широкими копытами, что необходимо для добывания корма (ягеля) из под снега. Т.е. в процессе эволюции выживают только те особи, которые обладают двумя описанными выше признаками (сильные ноги и широкие копыта).
Возникшие полезные признаки закрепляются у организмов за счет выживания тех особей, у которых имеются эти признаки и вымирания тех особей, у которых такие признаки отсутствуют.
В естественном отборе выделяют половой отбор , который представляет собой конкуренцию самцов за возможность размножения. Этой цели служит пение, ухаживание, брачный наряд, демонстративное поведение.
Помимо естественного, есть искусственный отбор , который осуществляется целенаправленно человеком. Результатом искусственного отбора являются новые породы животных, сорта растений и штаммы микроорганизмов с полезными для хозяйственной деятельности человека признаками.
Естественный отбор является важнейшей движущей силой эволюции и реализуется через борьбу за существование .
Борьба за существование выживание организмов, наилучшим образом приспособленных к данным конкретным условиям среды своего обитания называется борьбой за существование, является средством осуществления естественного отбора.
Ч. Дарвин выделил три формы борьбы за существование: внутривидовая, межвидовая и борьба с неблагоприятными условиями существования. Внутри- и межвидовая борьба основаны на конкуренции живых организмов за ресурсы (вода, пища, самки, места обитания и т.д.) и возможность оставить полноценное, плодовитое потомство.
Внутривидовая борьба является самым жестоким видом борьбы, т.к. организмы внутри вида имеют сходные предпочтения, а значит и более жесткую конкуренцию. Особенно ярко внутривидовая борьба проявляется среди животных. Так, среди хищных животных более сильные особи получают более полноценную пищу и в большем количестве. Это позволяет им выдержать конкурентную борьбу за самку и дать полноценное потомство, которому будут переданы признаки родителей. У павлинов большую вероятность оставить потомство будут иметь те особи, которые обладают наибольшим размером и красотой хвоста.
Межвидовая борьба за существование возникает между особями разных видов, занимающих одну экологическую нишу (живут на одной территории, питаются одними и теми же животными, для растений это борьба за свет, территорию и влагу). Сосна (светолюбивое растение) и ель (тенелюбивое) часто вступают в конкурентные взаимоотношения: семена ели легко прорастают под пологом соснового леса, но когда ель перерастает сосну, то сосна испытывает угнетение из-за затенения. Львы и волки (хищники), живущие в саванне на одной территории, питаются копытными и конкурируют из-за пищи.
В результате межвидовой борьбы у организмов разных видов возникают приспособления, позволяющие им занять разные экологические ниши и за счет этого существовать в более комфортных условиях . Так, жираф и зебра питаются одинаковой растительной пищей древесная растительность. Но они не конкурируют между собой, так как жирафы питаются листвой кроны деревьев, а зебры поверхностной растительностью.
Борьба с неблагоприятными условиями это выживание организмов в жестких неблагоприятных условиях существования (см. тему Гомеостаз и адаптации). Причиной возникновения приспособлений к условиям среды обитания является мутационная изменчивость , возникающая под влиянием условий окружающей среды. Возникшие мутации в случае их полезности закрепляются в потомстве за счет лучшего выживания особей, обладающих этими признаками и реализуются в форме приспособлений (адаптаций) .
Адаптации всегда носят относительный характер . Организмы, имеющие приспособления, полезные в одних условиях, оказываются совершенно неадаптированными в других, изменившихся условиях среды. Если, например, зеленого кузнечика перенести из зеленой травы в выгоревшую желтую, то покровительственная окраска больше не скрывает его, а наоборот, делает заметным для врагов.
Существует несколько разновидностей приспособленности организмов:
1) покровительственная окраска окрас, позволяющий организму быть незаметным на фоне окружающей среды. Примеры: зеленая окраска тли на фоне зеленых листьев капусты; темная окраска спины рыбы на темном фоне при взгляде сверху и светлая окраска брюха на светлом фоне при взгляде снизу; рыбы, живущие в зарослях водной растительности имеют полосатую окраску (щука) и т.д.
2) мимикрия и маскировка. Мимикрия состоит в том, что организм по форме похож на другой организм. Примером мимикрии является муха осовидка, форма ее тела напоминает осу и этим предостерегает от опасности, которой нет, так как эта муха не имеет жала. Маскировка состоит в том, что организм приобретает форму какого-то предмета окружающей среды и становится незаметным. Примером могут служить палочники - насекомые, по форме напоминающие обломки стеблей растений; есть насекомые, имеющие листообразную форму и т.д.
Рис. 86. Гусеницы пядениц в движении (вверху) и защитной позе (внизу слева)

Рис. 87. Кузнечик, крылья которого напоминают лист
3) предупреждающая окраска - организмы имеют яркую окраску, предупреждающую об опасности. Примеры: окраска ядовитых божьих коровок, пчел, ос, шмелей и т.д.
4) особые приспособления растений для реализации процессов опыления. Ветроопыляемые растения имеют длинные, свисающие тычинки, удлиненные, торчащие в разные стороны рыльца пестиков с приспособлениями для улавливания пыльцы и другие формы. Насекомоопыляемые растения имеют соцветия, яркую краску и экзотические формы цветка для привлечения определенного вида насекомого, с помощью которого реализуется опыление.
5) Особые формы поведения животных угрожающие позы и звуки, демонстративное поведение, зарывание страусом головы в песок и т.д.
Подводя итог, можно отметить, что результатом действия естественного отбора является развитие адаптаций и видообразование.
Видообразование это процесс разделения во времени и пространстве единого прежде вида на два или несколько самостоятельных новых видов. Возникает под влиянием приведенных выше эволюционных факторов в результате устойчивых изменений генотипической структуры популяций.
Представление о путях эволюционных преобразований это представления о путях достижения биологического прогресса или о главных направлениях эволюции. В современном виде сформулированы А.Н. Северцовым (1866-1936). Выделяют три главных направления в эволюции , каждое из которых ведет к биологическому прогрессу: ароморфоз (морфофизиологический прогресс), идиоадаптацию, общую дегенерацию.
Ароморфоз (от греч. «айро» поднимают, «морфа» форма) означает усложнение организации, поднятие ее на более высокий уровень. Изменения в строении животных в результате ароморфоза не являются приспособлениями к каким-либо специальным условиям среды, они носят общий характер и дают возможность расширить использование условий внешней среды (новые источники пищи, новые места обитания).
Ароморфозы животных обеспечивают переход от пассивного питания к активному (появление челюстей у позвоночных), повышают подвижность животных (появление скелета как места прикрепления мышц и замена пластов гладкой мускулатуры у червей на пучки поперечнополосатой у членистоногих), дыхательную функцию (возникновение жабр и легких), снабжение тканей кислородом (появление сердца у рыб и разделение артериального и венозного кровотока у птиц и млекопитающих).
К ароморфозам растений относится возникновение фотосинтезирующих организмов из гетеротрофов; возникновение псилофитов из водорослей; возникновение покрытосеменных с наличием двойного оплодотворения и новых оболочек у семени из голосеменных.
Общая черта ароморфозов заключается в том, что они сохраняются при дальнейшей эволюции и приводят к возникновению новых крупных систематических групп классов, типов, некоторых отрядов (у млекопитающих). После возникновения ароморфозов и особенно при выходе группы животных в новую среду обитания начинается приспособление отдельных популяций к условиям существования путем приобретения идиоадаптаций.
Идиоадаптация (от греч. «идиос» особенность, «адаптация» приспособление) приспособление к специальным условиям среды, полезное в борьбе за существование, но не изменяющее уровня организации. К идиоадаптациям относятся покровительственная окраска животных, колючки растений, плоская форма тела скатов и камбалы. К идиоадаптации можно отнести появление разных видов вьюрков на Галапогосских островах, различных грызунов, живущих в разных условиях (зайцы, белки, суслики, мышевидные грызуны) и другие примеры. В зависимости от условий обитания и образа жизни многочисленным преобразованиям подвергается пятипалая конечность млекопитающих.

Рис. 88. Разнообразие форм клюва у птиц обусловлено приспособлением к различной пище, т. е. является идиодаптацией: 1 зеленоклювый тукан, 2 колпица, 3 большеклювый попугай, 4 тупик, 5 фламинго, 6 водорез, 7 большой пестрый дятел, 8 большой кроншнеп, 9 утка, 10 шилоклювка, 11 ибис

Рис. 89. Общий вид бычьего цепня
Кроме основных направлений эволюции выделяют морфологические закономерности биологической эволюции : дивергенцию и конвергенцию.
Дивергенция это процесс расхождения признаков, в результате которого появляются новые виды, или возникшие в процессе эволюции виды отличаются друг от друга различными признаками, за счет приспособления к разным условиям существования. В результате дивергенции происходит появление новых видов из исходного вида.
Как указывал Дарвин, дивергенция лежит в основе всего эволюционного процесса. Дивергировать могут не только виды, но и роды, семейства, отряды. Дивергенция любого масштаба есть результат действия естественного отбора, т.к. сохраняются или устраняются виды (роды, семейства и т.д.) наиболее приспособленные к данным условиям. В результате дивергенции возникают гомологичные органы (см. выше).
Конвергенция (схождение признаков) выражается во внешнем сходстве организмов, обитающих в сходных условиях жизни. Так в одинаковых условиях существования животные, относящиеся к разным систематическим группам, могут приобретать сходное строение. Такое сходство строения возникает при сходстве функций и ограничивается лишь органами, непосредственно связанными с одними и теми же факторами среды (рис. 90).
Органы, образующиеся в результате конвергенции называются аналогичными (см. выше). Например, внешне очень похожи хамелеоны и лазающие агамы, обитающие на ветвях деревьев, хотя относятся к разным подотрядам. У позвоночных животных конвергентное сходство обнаруживают конечности морских рептилий и млекопитающих, жабры рака и рыбы, роющие конечности крота и медведки

Рис. 90. Конвергенция: развитие приспособлений для парения в воздухе
у позвоночных