Мезозойская эра, или эра средней жизни - довольно продолжительный отрезок геологической истории Земли длительностью около 175 млн. лет. Несмотря на то что мезозой затерян в дебрях далекого прошлого, о существовании в то время природных условий, разнообразных ландшафтов, о составе атмосферы, солености и глубинах морей и океанов, холодных и теплых течениях, животных и растительном покрове можно судить - уверенно и подробнее реконструировать их, чем например палеозоя. Отложения морского и континентального происхождения мезозойского возраста весьма многочисленны и распространены достаточно широко на всех континентах. Они содержат обильные остатки беспозвоночных и позвоночных животных, стволов и веток, отпечатки побегов и листьев, многочисленные споры и пыльцу растений. В отложениях заечатлены следы тектонических движений, вулканизма и экзогенных преобразований.
В отличие от более древних периодов геологического прошлого, когда о ландшафтах и особенностях органического мира мы могли судить только по изучению отложений современных континентов, начиная со среднего мезозоя появляются геологические данные по морям и океанам. Благодаря глубоководному бурению на основании изучения керна скважин получены неоценимые сведения не только о составе и возрасте пород залегающих на глубинах нескольких сот и даже тысяч метров от поверхности дна океанов, но и об условиях их образования.
В течение мезозоя в разных районах планеты периодически происходили интенсивные складкообразовательные и горообразовательные движения, сопровождаемые обильными вулканическими извержениями. Осуществлялось не только разрастание устойчивых областей земной коры, но и расколы континентов по крупным разломам с образованием рифтовых зон и активные перемещения литосферных плит.
Триасовый период - время раскола земной коры
В течение триасового периода, продолжавшегося около 35 млн. лет, в результате крупнейших расколов земной коры Пангея - этот супергигантский материк - разделился на два огромнейших материка - Гондвану и Лавразию.
Ранний триас - время полного завершения герцинского орогенеза. Поднятия и складкообразовательные движения в это время обозначились в Аппалачах, в пределах Мексиканского залива, на о-ве Тасмания, в Восточной Австралии, на юге Африки (Капские горы) и в Монголо-Охотском поясе.
Поднятиями были охвачены обширные платформенные участки. Области активизации платформ и орогенные области характеризовались не только активным воздыманием, но и формированием глубинных разломов и интенсивным вулканизмом. В триасовом периоде произошел распад Гондваны и Лавразии. Вдоль возникших глубинных разломов, предопределивших контуры современных материков южного полушария, впоследствии образовались глубокие протяженные впадины - рифтовые зоны, вначале весьма похожие на современные рифты Байкала или зону Великих Африканских разломов.
На границе палеозоя и мезозоя разломы рассекли многие устойчивые участки земной коры. Например, на Сибирской платформе они явились центрами излияний базальтовых лав, известных под названием траппов . Эти траппы имеют мощность около 15 км и занимают территорию свыше 500 тыс. км 2 .
В конце среднего триаса отдельные участки Тихоокеанского, Верхоянского, Кордильерского, Средиземноморского геосинклинальных поясов, а также некоторые герцинские структуры (Шпицберген, Мангышлак, Туаркыр), залитые морем, были приподняты и подверглись размыву.
В конце триасового периода произошла новая вспышка вулканизма. По ожившим древним и вновь образованным разломам на земную поверхность с глубины в несколько сот километров устремились потоки базальтовых лав. Под тяжестью многокилометровых толщ базальтов земная кора прогибалась, и по возникшим трещинам изливались новые порции базальтовых лав. Новая тенденция в развитии литосферы Земли наметилась в связи с расколом Пангеи и раздвижением континентов в разные стороны. В это время полностью закрылся океан палео-Тетис, Китайский континент причленился к Евразии, возник новый мезозойский океан Тетис, располагавшийся между Евразиатским и Африканским континентами. Тетис сильно сужался к западу и расширялся в восточном направлении. Это дало основание установить существование в Тетисе двух расходившихся на восток ветвей срединно-океанических хребтов. Северная ветвь охватывала Альпы, Карпаты, Балканы, Малый Кавказ и Эльбурс, а южная - Аппенины, Динариды, Анатолию и Южный Иран. Между этими хребтами располагались микроконтиненты: Итальянский, Родопский, Иранский. В то далекое время максимальная ширина мезозойского океана Тетис составляла 2500 км.
Вдоль рифтовых зон начала раскрываться Северная Атлантика, находившаяся между Северо-Американским и Африканским континентами. Южная Америка соединялась с Африкой, но отделялась от Северной Америки океаническими просторами, существовавшими на месте современного Карибского моря и Мексиканского залива.
В триасовом периоде произошло образование частей современного Северного Ледовитого океана. Во всяком случае, в настоящее время вскрыты погребенные под чехлом мощных толщ осадочных образований грабены, многие из которых располагаются в пределах Западно-Сибирской низменности и раскрываются в северном направлении. Западно-Сибирская низменность должна была стать океаном, но дальнейшее сближение Европы и Сибири помешало этому.
Растяжение литосферы с образованием океанов компенсировалось сжатием на континентальных окраинах, в результате которого появились вулкано-плутонические комплексы на окраине Евразии (Монголия, Забайкалье, Юго-Восточный Китай). На западных окраинах Северной и Южной Америки существовали протяженные островные дуги, сходные с современными островными дугами восточной акватории Тихого океана. Островные дуги имелись также в пределах современной Новой Зеландии и Новой Каледонии, а также в Западной Антарктиде.
В тесной зависимости от тектонической жизни планеты развивались морские трансгрессии и регрессии. Рост размеров суши и увеличение ее гипсометрического уровня в основном осуществлялся за счет горообразовательных процессов, проявлявшихся на границах литосферных плит.
В среднем триасе сократились площади осадконакопления на платформах, и многие из них были приподняты и подвержены размыву. На окраинах и в центральных частях древних платформ и эпигерцинских плит образовались узкие, глубокие грабенообразные прогибы - авлакогены, которые заполнялись грубым терригенным материалом, а иногда и угленосными толщами.
В позднетриасовую эпоху площадь эпиконтинентальных морей увеличилась незначительно, однако трансгрессии были кратковременными. В это время возникли складчатые структуры, так называемые ранние мезозоиды . Древнекиммерийская фаза складчатости проявилась в Индонезии, Индокитае, Тибете, Северном Афганистану, на Эльбурсе, Кавказе и Балканах.
Однако несмотря на довольно обширные поднятия, рельеф континентов не отличается большими контрастами и высотой. На платформах господствовали аккумулятивные и денудационные равнины. К первым относятся крупные озерно-аллювиальные внутриконтинентальные комплексы, озерно-дельтовые приморские и эоловые (пустынные) низменности с небольшими увалистыми водораздельными возвышенностями, а ко вторым - денудационные холмистые и возвышенные равнины. Спокойные плавные очертания рельефа иногда нарушались узкими и протяженными впадинами. В них обычно располагались речные долины и озера. Глубина морей, покрывавших современные континенты, была небольшой. Только в геосинклинальных областях и вблизи срединно-океанических хребтов существовал контрастный рельеф с перепадами глубин в несколько тысяч метров.
Сильные изменения произошли в триасовом периоде в составе органического мира. На смену вымершим в конце перми группам фауны и флоры пришли новые. Наземный растительный мир знаменовался появлением важнейших представителей хвойных и дальнейшим развитием гинкговых и цикадофитовых. Наряду с ними продолжали существовать реликты палеозойских семенных папоротников, кордаитов и каламитов, которые, однако, уже в середине триаса вымерли.
Животному царству суши было свойственно преобладание рептилий и заметное сокращение земноводных, в частности стегоцефалов. Существенные изменения произошли в классе рептилий. Зверообразные и котилозавры вымерли, и им на смену в середине триаса пришли динозавры. В триасе появились и первые млекопитающие. Это были мелкие и весьма немногочисленные животные.
Органический мир морей был довольно богат и многообразен. В мелководных, нормально-соленых условиях большого расцвета достигли головоногие моллюски - аммониты и белемниты. Аммониты в отличие от своих позднепалеозойских предков имели более сложное строение и весьма богатую и разнообразную скульптуру горизонтально завитой раковины. Сильно видоизменилась фауна двустворчатых моллюсков - появились устрицы, возникли новые роды среди брюхоногих моллюсков и морских ежей, а также первые представители шестилучевых кораллов. Большого разнообразия достигли морские позвоночные. К плавающим пресмыкающимся относились многочисленные ихтиозавры и плезиозавры, а среди рыб преобладали костистые. Практически повсеместное распространение имел однотипный теплолюбивый комплекс морских беспозвоночных. Например, остатки одних и тех же рептилий и амфибий, жизнедеятельность которых прекращалась при среднегодовых температурах ниже 10°С, обнаружены даже на крайнем северо-востоке и востоке Евразии и в южной части Австралии, т. е. в тех районах, которые по палеомагнитным данным располагались вблизи Северного и Южного полюсов. По-видимому, даже в околополярных районах в раннем и среднем триасе температура вряд ли опускалась ниже уровня, характерного для субтропического пояса.
В течение раннего и среднего триаса на Земле существовали четыре основные природные зоны: экстрааридная (пустынная), умеренно аридная зона сухих и опустыненных саванн, переменно-влажная и равномерно-влажная.
Экстрааридные условия господствовали на значительной части Северо-Американского, Африканского, Евразиатского материков, на севере Южной Америки и в Австралии. В этих районах располагались крупные пустыни с характерными эоловыми формами рельефа - дюнами, барханами и такырами. Пустыни пересекались только редкими транзитными крупными реками. В одиночных оазисах, находившихся на берегах водоемов и в речных долинах, произрастали устойчивые к засухе деревья, главным образом своеобразные хвойные и цикадофитовые. В редких пресноводных и слабозасолоненных бассейнах существовал резко обедненный в видовом и родовом отношении комплекс остракод и двоякодышащих рыб.
Более умеренные аридные тропические условия в течение значительной части триаса преобладали на севере Северной и Южной Америки, в Северной Европе, Западной и Южной Сибири, Китае и Монголии. Растительный покров не был сплошным, но оазисы встречались значительно чаще, чем в экстрааридной зоне. Состав растительности в них был довольно разнообразный. Наряду с позднепалеозойскими реликтами (папоротники и кордовые) здесь господствовали хвощевые.
Густые заросли из сравнительно влаголюбивых ассоциаций растений располагались в широких долинах, на побережьях крупных озер и приморских низменностях.
В переменно-влажной области тропического климата господствующим типом ландшафта являлась лесная саванна. Фауна морей переменно-влажной и умеренно аридной областей тропического пояса отличалась богатством видов и родов. На шельфах наряду с кораллами, брюхоногими двустворчатыми и головоногими моллюсками, мшанками, морскими ежами и лилиями обитали рыбы и водные рептилии. Среди последних особо выделялись ихтиозавры (рыбоящеры) и плезиозавры. Ихтиозавры - эти хищники мезозойских морей - были великолепными пловцами. Размеры их превышали 2 м. Они имели гладкую кожу, удлиненный череп с зубастой пастью. Тело заканчивалось вертикальным кожистым хвостовым плавником. В отличие от ихтиозавров плезиозавры имели массивное тело, короткий хвост, ластообразные конечности, очень длинную шею и маленькую голову. Массивность и строение тела позволяли плезиозаврам обитать только в воде и передвигаться в пределах обширных мелководий с глубинами, не превышающими 2-4 м. Их размеры достигали 15 м.
Приморские и внутриматериковые озерно-аллювиальные равнины в областях переменно-влажного и умеренно аридного климата являлись основным местообитанием рептилий. В этих условиях существовали ящеротазовые динозавры (их иногда называют ужасными ящерами ) и амфибии.
Большое распространение в триасе имели листрозаурусы . Эти пресмыкающиеся достигали в длину 120 см. Массивное тело держалось на коротких и сильных ногах, челюсть изгибалась кверху, все зубы были направлены внутрь и только два небольших клыка выпирали наружу. Находки скелетов листрозаурусов, этих типично сухопутных животных, в Африке, Южной Америке, Индии, Австралии и Антарктиде доказывают единство материков в виде Гондваны и отсутствие в триасовое время каких-либо разделявших их крупных водных преград.
Равномерно-влажные тропические условия в течение всего триаса господствовали в Юго-Восточной Азии, Индии, Южной Америке и Антарктиде. Здесь главенствующее положение занимали лесные и озерно-болотные ландшафты. В составе лесов принимали участие такие представители голосеменных, как цикадофитовые, беннеттитовые, гинкговые, а также древовидные папоротники, плауны и хвощи.
На приморских озерно-дельтовых и озерно-аллювиальных низменностях в условиях экваториального влажного климата располагались густые заросли лесов, напоминавших современные дождевые экваториальные леса. В их состав входили древовидные папоротники, цикадофитовые и беннетитовые. Обилие влаги и растительной массы способствовало накоплению залежей угля.
Хотя площадь аридного климата сократилась, но особенности осадконакопления и характер жизнедеятельности животных по сравнению по сравнению со средним триасом существенно не изменились.
Наиболее прохладные условия в течение триасового периода существовали на северо-востоке Евразии. По сравнению с другими областями здесь обнаружено очень малое содержание карбонатного материала (одного из основных показателей теплого климата), установлен иной тип растительности и животного царства, в котором отсутствуют типичные группы тропических организмов (кораллы, морские ежи). Остатки морских беспозвоночных обладают нежными и тонкими раковинами.
На протяжении триасового периода наблюдается очень интересная зависимость организмов от окружающей среды. Вследствие возрастания сухости воздуха в первой половине триаса многие рептилии были вынуждены проводить значительную часть времени в неглубоких озерах, реках и в прибрежных зонах морей. Причем некоторые из них (ихтиозавры и плезиозавры) по мере увеличения аридизации стали совсем водными формами. Другие, наоборот, приспособились к обитанию на суше и приобрели толстый роговой панцирь, предохраняющий их от испарения и перегревания. В конце триаса ввиду сильного изменения ландшафтно-климатических обстановок произошло обновление фауны рептилий.
Юрский период - расцвет динозавров
Юрский период - один из самых характерных периодов мезозойской эры. Во многих отношениях он отличается от триасового, и главное заключается в том, что юра - это время типично морских условий в жизни континентов. Отмечаются две морские трансгрессии, которые одновременно охватили почти все континенты. Они проявились в конце раннеюрской и начале позднеюрской эпохи и разделялись отчетливой регрессией среднеюрской эпохи.
Тектоническая жизнь на планете в течение юрского периода была более интенсивной, чем в триасе. Для этого периода типичны процессы складко- и горообразования, происходившие в результате столкновения литосферных плит, и крупные нарушения глыбового характера в пределах платформ и областей активизации. Образование горстовых поднятий и грабенов сопровождалось интенсивным наземным вулканизмом. Древнекиммерийский орогенез затронул отдельные районы Альпийско-Гималайского пояса (Пиренеи, Альпы, Карпаты, Крым, Кавказ, Тибет, Индокитай). Орогенические движения происходили в конце средней юры, но особой силы они достигли в позднеюрскую эпоху, когда охватили северные районы Тихоокеанской геосинклинального пояса.

В это же время Австралия и Антарктида отделились от Африки и Индии, и возникший новый Индийский океан соединился с восточной окраиной океана Тетис. Наиболее обширным в юрском периоде был Тихий океан. Его размеры даже превышали современные. Азия отделилась от Северной Америки океаническим бассейном. На западной окраине Северной Америки происходила интенсивная складчатость - так называемый невадийский орогенез, который по времени соответствовал позднекиммерийскому. Складкообразование сопровождалось внедрением крупных интрузий и высокотемпературным метаморфизмом. Образование невадийского пояса складчатости и батолитов было обусловлено процессом столкновения континента с островной дугой.
Активная тектоническая жизнь непосредственно оказывала влияние на распределение морских бассейнов и рельеф земной поверхности. В начале поздней юры наступила одна из крупнейших мезозойских трансгрессий. В это время морские воды проникали по вновь образованным грабенообразным структурам далеко в глубь платформ. Море затопило не только значительную часть Евразии, Северной Америки, Южной Америки, но и восточное и западное побережье современной Африки, западное побережье Мадагаскара, Индию и Австралию. Однако в результате активизации тектонических движений в конце юрского периода в ряде районов наступила регрессия моря. Это обусловило не только обмеление морских бассейнов, но и возникновение обширных лагун и заливов с повышенной соленостью воды.
Различная интенсивность тектонических движений привела к формированию разнообразных крупных форм рельефа земной поверхности. Максимальная глубина вновь образованных океанических бассейнов не превышала нескольких тысяч метров. В их пределах выделялись абиссальные и батиальные области, зоны материкового склона и обширные шельфовые участки, постепенно переходящие в эпиконтинентальные моря. Максимальная глубина последних составляла 400-500 м.
Наиболее сложным и контрастным рельефом обладали области, испытавшие киммерийские движения. Для них были характерны неровное дно морских бассейнов, соседство глубоководных впадин с поднятыми участками дна и островными дугами, расчлененное побережье и сильно изрезанная береговая линия. Приморские низменности сравнительно быстро переходили в возвышенности и горные массивы, которые прорезывались многочисленными реками. Менее контрастный рельеф существовал на платформах. Лишь в центральных их частях располагались выровненные возвышенности и холмисто-увалистые равнины сменяющиеся аккумулятивными озерно-аллювиальными и озерно-дельтовыми приморскими низменностями.
Юрская флора характеризовалась господством голосеменных форм. Споровые, в основном папоротниковые и хвощевые образовывали самостоятельные заросли в пределах отдельных сильно увлажненных низменностей, примыкавших к морям и крупным озерам. Растительный покров в первой половине юрского периода был представлен хвойными, цикадофитовыми, беннеттитовыми и гинкговыми. В поздней юре в связи с снижением влажности леса поредели, исчезли влаголюбивые формы и появились растения, способные переносить засуху. Вначале редколесья располагались на возвышенностях, но впоследствии из-за роста засушливости они распространились и на приморские низменности.
Флора юрского периода была хорошо дифференцирована как в экологическом, так и в географическом плане. Это определялось воздействием зонального и провинциального климата высотой суши и соотношением площади суши и моря, наличием крупных орографических рубежей и т. д.
По экологическому характеру и систематическому составу в течение ранней и средней юры на платформах северного полушария, флора которых в настоящее время хорошо изучена существовали, по крайней мере, три геоботанические зоны Флора, произраставшая на северо-востоке Евразии и северо-западе Северной Америки, обладала наименьшим систематическим разнообразием и характеризовалась слабым развитием теплолюбивых форм и отсутствием представителей тропической и экваториальной флоры. Здесь росли главным образом хвойно-гинкговые леса.
Теплолюбивые цикадофитовые и беннеттитовые появились только на южных окраинах зоны хвойно-гинкговых лесов. По сравнению со своими сородичами из южных областей они имели небольшую высоту и слабо разветвленный ствол. Роль травянистого покрова в хвойно-гинкговых лесах играли низкорослые папоротники, мелкие хвощи и плауны. Деревья обладали годовыми кольцами нарастания, подобными тем, которые имеются современных деревьев, растущих в условиях сезонных колебаний температур. Однако, по мнению известного советского палеоботаника В. А. Вахрамеева, климат зоны хвойно-гинкговых лесов вряд ли можно было считать суровым. Несмотря на хорошо выраженную сезонность климата, зимы были мягкими: средние зимние температуры не опускались ниже 0°С. Многие гинкговые в прохладные сезоны года теряли листву.
В юрское время проявилась вертикальная климатическая зональность, обусловленная существованием значительной разницы в температурном режиме и увлажнении между низменостями, возвышенностями и горными массивами. В горах произрастали хвойные леса, а на хорошо дренированных низменностях - хвойно-гинкговые и гинкговые леса. В областях высоким температурным режимом на низменностях росли цикадофитовые, беннеттитовые и смешанные - цикадофитово-беннеттитовые леса, а на слабо дренированных низменностях, подвергавшихся заболачиванию, и на широких заболоченных поймах располагались заросли папоротников и хвощей.
В ранней и средней юре в зоне хвойно-гинкговых лесов существовала необычайно высокая увлажненность, что и предопределило развитие обильно увлажненных ландшафтов. Здесь располагались озерно-болотные и сильно заболоченные низменности. В широких речных долинах находились пойменные заболачивающиеся озера и старицы. В них, а также в дельтах полноводных рек накопление растительных остатков способствовало формированию угленосных осадков.
Наибольшее разнообразие имела тропическая флора, в которой преобладали цикадофитовые и беннеттитовые. Исходя из разного систематического состава в различных ландшафтах тропического пояса выделяются несколько палеофлористических областей и зон. Одной из многих являлась Индо-Европейская область, расположенная в пределах Евразиатского материка. Примерно такая же флора произрастала на Тихоокеанском побережье Северной Америки, в Австралии и в Южной Америке. Значительные изменения в составе растительного покрова и географическом распространении различных форм произошли в позднеюрскую эпоху. Только флора южных и северных приполярных районов по существу сохранила свой прежний облик. Благодаря высокой влажности и сохранившемуся температурному режиму здесь продолжали рост хвойно-гинкговые леса с папоротниковым подлеском. Постепенно смещавшаяся в северном полушарии засушливость климата вносила определенные коррективы не только в распределение ландшафтных обстановок, но и в состав и распределение растительного покрова. В районах, которые подверглись сильной аридизации, многие влаголюбивые формы исчезли, леса поредели, и в них сохранились лишь засухоустойчивые формы. Практически полностью исчезли гинкговые и папоротниковые. В целом растительный покров в областях умеренного увлажнения стал напоминать в экологическом отношении современные леса восточной части Средиземноморья. Растительность ксерофильного редколесья обладала чертами, свойственными засушливому климату. Беннеттитовые и цикадофитовые имели кожистые листья, а хвойные – чешуйчатую хвою.
В низких широтах из-за высокой степени аридизации растительный покров был сильно разрежен, а местами полностью отсутствовал. Деревья произрастали только в долинах рек и вблизи источников воды. Здесь основная роль принадлежала опустыненным саваннам, саванно-степям и полупустыням. В областях с сильным дефицитом влаги располагались пустыни.
В низких широтах, в областях обильного увлажнения, тип растительного покрова в поздней юре по сравнению с ранне- и среднеюрской эпохами почти изменился. Здесь произрастали влаголюбивые тропические и экваториальные сообщества бенеттиттовых, цикадофитов и древовидных папоротников.
Фауна позвоночных юрского периода характеризовалась значительным развитием гигантских рептилий, среди которых различаются наземные, водные и летающие формы, травоядные и хищные.
Родоначальником динозавров являлись сравнительно широко распространенные еще в конце триаса мелкие ящеры – текодонты. В триасе эти ящеры стали передвигаться на двух задних ногах. Поскольку главным органом чувств у рептилий были глаза, преимуществом по сравнению с другими обладали особи с высоко поднятой головой. Они лучше ориентировались среди высокой травянистой растительности.
Многие из динозавров имели гигантские размеры. Длина некотороых из них была 20-25 м, а масса достигала 30-35 т. Среди динозавров были распространены как четырехногие формы, так и двуногие. Многие гиганты вели полуназемный образ жизни. Значительную часть времени они, подобно современным бегемотам, проводили в воде. Для поддержания большого веса тела кости скелета должны были обладать значительной прочностью. Однако быстро перемещаться по суше такие гиганты не могли. Они обладали полыми костями скелета, тонкими зубами, пригодными для поедания только мягких побегов и водорослей, и облегченными шейными позвонками. Динозавры, ведущие полуназемный образ жизни, имели далеко отодвинутые глаза и длинные носовые (воздухоносные) ходы. Благодаря этому, например, утконосые динозавры зауролофы могли полностью погружаться в воду, спасаясь от врагов или в поисках пищи.
Некоторые полуназемные динозавры имели укороченные передние конечности, снабженные перепонками, длинный уплощенный хвост. Они были хорошими пловцами. В то время как полуназемные динозавры передвигались преимущественно на четырех ногах (диплодоки, бронтозавры) и реже на двух (зауролофы), типично наземные формы, за малым исключением, были двуногими. Они обитали главным образом в сильно увлажненных местах, покрытых богатой растительностью, но с твердой почвой, так как в сильно заболоченных или густозалесенных местах Нои не смогли бы передвигаться. Среди наземных динозавров различались растительноядные и хищные. Ярким представителем растительноядных динозавров были игуанодонты . Некоторые из них достигали в длину 10 м. Эти животные имели трехпалые задние конечности с перепонками, которые позволяли перемещаться по топким побережьям рек, озер и морей. Они обладали своеобразным клювом, который использовался при объедании побегов с кустарников и деревьев и как средство защиты. Животное имело сильную челюсть и могло наносить клювом опасные укусы и рваные раны. Страшными хищниками, жившими в юрском периоде, были крупные двуногие динозавры - кариозавры .
Наземные динозавры размножались с помощью яиц, которые откладывали в теплый песок, точно так же как это в настоящее время делают крокодилы и черепахи. Растительноядные динозавры вели стадный образ жизни. Об этом свидетельствует множество отпечатков следов, обнаруженных в ископаемом состоянии на берегах древних морей и крупных озер.
Обилие сильно заболоченных территорий, густая лесная растительность, наличие многоводных и широких рек, крупных озерных бассейнов в ранней и средней юре способствовали значительному сокращению численности наземной динозавровой фауны и смещению ареала их обитания в пределы ксерофильного редколесья. Господствующее положение в сильно обводненных ландшафтах заняли полуназемные и водные рептилии. Среди последних главенствовали ихтиозавры и плезиозавры. Ихтиозавры своей внешностью напоминали современных дельфинов, но в отличие от них имели более удлиненную морду с зубастой пастью. Длина их тела достигала 6 м. Вместо передних и задних ног у них были плавники в форме ласт. Ихтиозавры, так же как и млекопитающие, были живородящими животными.
Современниками ихтиозавров были плезиозавры, плиозавры и др. Плезиозавры и плиозавры обладали мощными ластами. Они довольно свободно чувствовали себя в воде. Небольшая голова с зубастой пастью, в которой находилось множество острых конических зубов, располагалась на очень длинной шее. Наиболее крупные особи достигали в длину 10 м.
Постепенно ряд рептилий приспособился к обитанию в воздушной среде. Предками юрских летающих ящеров - птерозавров были текодонты, а точнее, те из них, которые вели древесный образ жизни и обладали способностью планировать с ветки на ветку, подобно некоторым современным формам Планирование на небольшие расстояния у одних особей осуществлялось благодаря длинным чешуям которые расширялись на концах, образуя тонкостенные мешочки, у других – с помощью эластичной кожистой пленки, натянутой между конечностями и туловищами.
Летающие ящеры (рамфоринхи, птеродактили и птеронодоны ) быстро освоили воздушный океан. Они были представлены многочисленными видами разных размеров от карликов до гигантов. Рамфоринхи имели длинный хвост, пятипалые задние конечности и длинные узкие перепончатые крылья размахом около 1 м. Гигантами среди летающих ящеров следует признать птеродактилей, многие из которых достигали в размахе крыльев около 8 м. Однако это, как стало известно, не предел, поскольку найдены скелеты, по которым можно судить, что отдельные особи имели размах крыльев около 15 м.
Челюсти летающих ящеров были густо усажены острыми и длинными зубами, тело одних покрывала густая шерсть, а тело других - чешуя, крылья были перепончатыми.
В юрский период появились первые птицы - археоптериксы и археорнисы, которые были величиной с голубя. Необходимо отметить, что эти птицы имели много общих черт с пресмыкающимися, особенно в строении черепа и скелета. Вместе с тем они обладали более совершенной кровеносной и нервной системой, у них были лучше развиты органы чувств по сравнению с рептилиями. Большой объем мозга, забота о потомстве и хорошая приспособляемость к различным условиям жизни способствовали широкому расселению птиц и обитанию в самых разнообразных ландшафтно-климатических условиях.
В пресных озерах, а также в старицах и полноводных реках обитала моллюсковая фауна. Озера, расположенные на низменностях, и широкие поймы рек зарастали водной и прибрежной растительностью и превращались в болота. Здесь происходило накопление мощных толщ угля. Воды озер и стариц, содержащие большое количество продуктов гниения растений, были бедны кислородом, известью и многими питательными веществами. Это привело к тому, что придонная фауна водоемов состояла из однообразных остракод и моллюсков.
В поздней юре в результате иссушения климата и исчезновения густой и влаголюбивой растительности многие водные бассейны из заболоченных превратились в открытые и лишенные растительности, что в свою очередь привело к увеличению в воде кислорода и извести. Господствующее положение заняли формы, экологически связанные с теплыми и прозрачными водами. Это различного типа крупные формы двустворчатых и брюхоногих моллюсков.
Богатством видов отличалась в юрском периоде морская фауна беспозвоночных. Причем в ее распространении намечается ряд различий, позволяющих наметить существование зоогеографических провинций и областей. Так, например, в пределах северного полушария выделяются по крайней мере три экологических типа фауны: бореальный (северный), смешанный и тропический. Наиболее богатой и разнообразной была фауна беспозвоночных тропических морей. В сторону полюса она сильно обеднялась.
В ранней и средней юре в тропических и бореальных морях обитали аммониты и белемниты, а также двустворчатые моллюски. Ряд типичных бореальных форм мигрировал в низкие широты, а в поздней юре наблюдалась обратная картина. В высокие широты перемещались тропические представители головоногих и двустворчатых моллюсков и брахиопод, кораллы, морские ежи и строматопоры.
Расцвет коралловой фауны произошел в середине позднеюрской эпохи. В это время существовало максимальное количество семейств и родов кораллов, а во многих морских бассейнах в зоне мелководья возникли многочисленные рифовые постройки. Крупнейшие рифовые тела существовали на перифериях тропического океана Тетис, в эпиконтинентальных тропических морях Северной и Южной Америки и Африки.
В течение юрского периода наблюдались значительные изменения климата. Раннеюрская эпоха характеризовалась ярко выраженной гумидизацией климата, большим разнообразием сильно обводненных ландшафтов, присутствием в осадочных образованиях растительного углерода, причем не только в рассеянном состоянии, но и в виде мощных толщ каменных и бурых углей. По температурному режиму в юрском периоде можно выделить экваториальный, два тропических и два субтропических пояса. В экваториальном и тропических поясах среднегодовые температуры обычно превышали 22-24°С.
В ранней и средней юре существовала не только термическая зональность, но и зональность, обусловленная различием во влажности и количестве выпадавших атмосферных осадков. Выделяются северный и южный аридные пояса и расположенный между ними экваториальный влажный пояс. К северному аридному поясу относились центральные районы США и Северная Африка, а к южному - Карибский бассейн, территория Сомали и Танзании. Значительно большими площадями обладали районы с переменным увлажнением - Африка, Аравия, юг Индокитая, юг Бразилии. Характерными ландшафтами перечисленных районов являлись ксерофильное редколесье, располагавшееся на берегах морей, озер и крупных речных артерий, а в аридных поясах - полупустыни и пустыни. Соленость воды морских бассейнов этих территорий не превышала 35 ‰ .
Районы, расположенные в средних широтах, а в северном полушарии такими были Западная и Восточная Сибирь, Северо-Восток СССР, Монголия и Северный Китай, характеризовались развитием влажного субтропического климата. Однако его главное отличие от современного субтропического климата заключалось в отсутствии четко выраженного зимнего сезона с отрицательными температурами. Условно северная граница субтропического климата в Евразии намечается по линии Белое море - Южный Урал - Балхаш - Шаньдунский полуостров. Растительным эквивалентом этого климатического пояса являлась зона хвойно-гинкговых лесов.
В этом климатическом поясе в основном располагались озера болота широкие поймы и обширные дельты рек, а также моря с нормальной соленостью. На низких водоразделах, в пределах увалисто-холмистых равнин и преимущественно на возвышенностях и склонах гор произрастали хвойные леса.
Присутствие в составе хвойно-гинкговых лесов значительного количества цикадофитов, распространение каолинитовых кор выветривания, наличие среди морской фауны рептилий и теплолюбивых моллюсков, примесь в морских осадках глауконита и стяжений фосфорита, а также палеотемпературные определения позволяют считать, что на севере субтропического пояса среднегодовые температуры колебались в пределах 15-20°, а на юге - 18-20 °С. Близкие условия существовали в средних и высоких широтах южного полушария. Температурные условия в среднеюрскую эпоху после кратковременного понижения на границе ранней и средней юры вновь стали высокими.
Несмотря на большую влажность, в результате продолжительного прохладного сезона выветривание не доходило до каолинитовой стадии. Морская фауна стала беднее за счет исчезновения теплолюбивых форм, а на континентальных пространствах произрастали хвойно-гинкговые и хвойные леса.
Иссушение климата в поздней юре привело к постепенному исчезновению лесной растительности, высыханию озер и к сокращению объема поверхностных вод. В условиях сильного дефицита влаги ландшафты приобрели вид современных опусты-ненных саванн, полупустынь и пустынь.
По степени распределения влажности в поздней юре в аридном поясе северного полушария выделяются две области. В центральной части аридного пояса существовали наиболее сухие условия с продолжительными засухами, а в периферических частях - на территории современной Центральной Европы, на юге СССР, в Монголии и Северном Китае периоды засухи были менее продолжительными. Это способствовало развитию ксерофильного редколесья, озерных водоемов и рек с обширными поймами. Ландшафтно-климатические особенности этой области, так же как и области переменно-влажного климата, в пределах которых располагались обширные низменности, покрытые разреженными лесами, с твердым грунтом на побережьях многочисленных неглубоких озер, весьма благоприятствовали развитию динозавровой фауны.
Субтропический пояс выделяется для территории Евразии, и по отношению к современным широтам он простирался субмеридионально. Субтропический климат господствовал в Северной Европе, на значительной части Восточно-Европейской платформы, на Урале, в Сибири, Монголии. Здесь среднегодовые температуры изменялись в пределах 12,5-13°С. Даже в бассейне Печоры, на Таймыре и на островах Земли Франца-Иосифа среднегодовые температуры морских вод в позднеюрскую эпоху временами достигали 17,5°С. Наиболее низкие температуры были характерны для умеренного пояса. Умеренный климат в это время господствовал в северном полушарии на Крайнем Северо-Востоке СССР и Аляске. В морях умеренного пояса обитала бореальная фауна, состоявшая главным образом из тонкостенных двустворча тых моллюсков и редких белемнитов.
Меловой период - становление океанов и возникновение цветковых
Мезозойская эра заканчивается меловым периодом, продолжавшимся около 70 млн. лет. Он знаменовал собой качественно новый этап в развитии ландшафтно-климатических областей и органического мира Земли. В течение этого периода тектоническая жизнь планеты оставалась активной.
В течение мелового периода продолжались киммерийские горообразовательные движения. В начале мелового периода в результате этих движений были созданы горно-складчатые сооружения на востоке Евразии. Складчатые пояса располагались на огромной территории, охватывающей Верхоянье, Чукотку, Камчатку, Дальний Восток и восток Китая. Дополнительные складкообразовательные движения на границе раннего и позднего мела (так называемая австрийская фаза складчатости) усилили контрастность этих участков земной коры. В это время, как и ранее, наряду с подводными вулканическими излияниями осуществлялось внедрение гранитных интрузий. Движения, обусловленные австрийской фазой складчатости, наиболее сильно проявились в Восточной Азии и Кордильерах и менее интенсивно в современном Альпийско-Гималайском поясе (Восточные Альпы, Карпаты, Кавказ, Памир). В конце мелового периода имела место довольно активная ларамийская орогеническая фаза. Складчатые движения охватили окраины Тихого океана. На значительную высоту оказались приподняты Анды, Кордильеры, горные сооружения Антарктиды, Чукотки и Камчатки.
Продолжалось раздвижение гондванских континентов. Северная Америка отделялась от Африки и Евразии. Эти активные раздвижения сопровождались формированием новых морей и океанов. Так, например, Атлантический океан приобрел близкие к современным очертания, возникли Бискайский залив и Гренландское море. В это время был заложен рифт Красного моря и началось раскрытие Индийского океана, а Индостан и Мадагаскар отделились от Африки. Индостанский континент совершил длительное путешествие, исчисляемое тысячами километров, пока в середине палеогена он не столкнулся с Азиатским континентом. В результате такого столкновения были образованы Гималаи.
Началу мелового периода был свойствен регрессивный этап развития, но в середине раннего мела ему на смену пришел трансгрессивный этап. Позднемеловая морская трансгрессия была одной из самых величайших трансгрессий на Земле. В это время многие континенты оказались покрытыми морем. Море занимало почти 40% территории современной Евразии и 35% территории Северной Америки, около 20-25% территории Африки и Австралии и 15% территории Южной Америки.
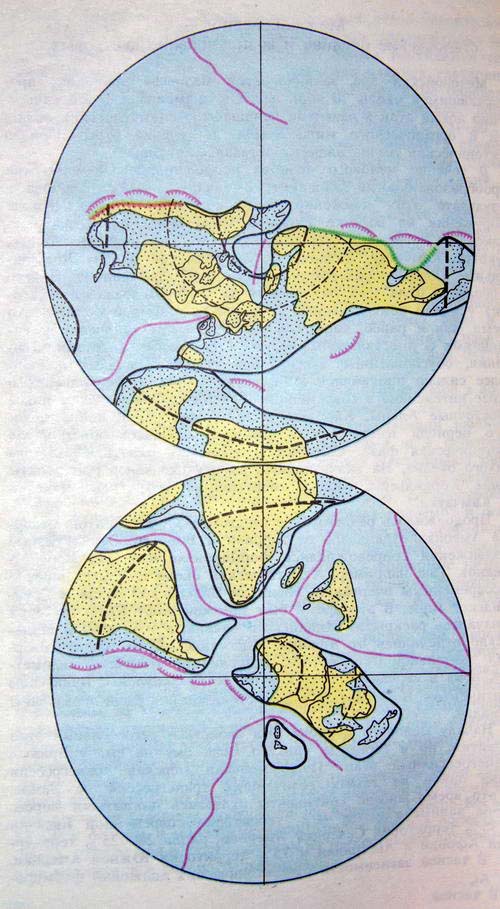
Положение литосферных плит и срединно-океанических хребтов в позднемеловой эпохе
В тесной зависимости от тектонических движений формировался рельеф земной поверхности. Наиболее контрастным и возвышенным он был в местах столкновения литосферных плит между собой. В результате поднятий и складчатости были образованы многие горные массивы. Области платформенной активизации также обладали возвышенным и горным рельефом.
Так же как и в настоящее время, горные сооружения и глубоководные океанические впадины занимали на земной поверхности весьма незначительную площадь. Основная же роль принадлежала выровненным шельфовым участкам, неглубоким эпиконтинентальным морям, приморским озерно-дельтовым и внутриконтинентальным озерно-аллювиальным низменностям и равнинам, возвышенным равнинам с увалисто-холмистыми водоразделами и выровненным плато. На них, а также на плоских водоразделах в благоприятных климатических условиях формировались коры выветривания. Некоторые более древние горные сооружения (герциниды, каледониды) в эпохи активизации испытали поднятия, однако амплитуда высот была не очень значительной и вряд ли превышала 1000 м.
В течение мелового периода развитие организмов и их расселение определялись природными условиями. Значительные изменения в распределении ландшафтно-климатических зон в первую очередь отразились на наземной флоре. В середине мелового периода осуществлялось широкое расселение многих представителей высшей флоры - покрытосеменных растений. Еще в начале мелового периода их роль в растительном покрове была невелика, но уже в середине позднемеловой эпохи они заняли главенствующее положение.
По сравнению с поздней юрой в раннем мелу произошло изменение морфоанатомического строения гинкговых. В то время как юрские гинкговые обладали сильно расчлененной листовой пластинкой, раннемеловые формы имели уже цельную пластину. Это свидетельствует не столько о более высокой ступени развития, сколько об их приспособлении к сезонным, а возможно, и к суточным колебаниям температур.
Многие древесные сообщества хвойных и гинкговых, особенно те, которые росли в высоких и средних широтах, обладали ясно выраженными годичными кольцами. Это свидетельствует о существовании в зоне хвойно-гинкговых лесов относительно прохладных условий.
В средних широтах располагалась зона цикадофитово-беннеттитовых лесов. Она отделялась от зоны хвойно-гинкговых лесов довольно узкой полосой развития смешанных цикадофитово-хвойно-гинкговых лесов. В Евразии к этой зоне относились крайний север Европы, Северный Урал, юг Западной Сибири и Приморье.
Наивысшего расцвета и разнообразия достигла флора в областях с тропическим влажным климатом. Склоны возвышенностей и низменности были покрыты густым пологом леса, весьма сходного с современными экваториальными лесами. Особенно большого разнообразия достигли цикадофитовые и многочисленные древовидные папоротники. В сторону областей засушливыми условиями многие влаголюбивые формы исчезали, лесной покров редел и превращался в ксерофильное редколесье.
Лесная растительность тропического типа произрастала Южной Евразии, на юге Северной Африки, юге Гренландии, также в Африке, Австралии и Южной Америке.
В начале раннемеловой эпохи в лесах, состоящих из голосеменных форм, появились первые представители покрытосеменных растений. Уже в конце раннего мела их роль резко увеличилась. Сильно уменьшился видовой состав голосеменных, несмотря на существование довольно благоприятных условий.
Появились и сравнительно быстро распространились по всему земному шару магнолии, лавровые, платаны, эвкалипты и дубы.
Среди хвойных господствующее положение заняли сосновые и таксодиевые. Эти две группы растений по условиям произрастания сильно отличались друг от друга. Сосновые леса, хорошо приспособленные к интенсивному солнечному облучению и засушливым условиям, произрастали на открытых возвышенных участках. Таксодиевые же, являющиеся тенелюбивыми растениями, предпочитали области с сильным увлажнением. Поэтому заросли таксодиевых располагались в низменных, обильно увлажненных местах, главным образом на низких поймах. В конце раннего мела постепенно исчезли многие мезозойские формы (подозамитовые и саговые) и сильно обновился состав папоротников, особенно среди травянистых форм.
В раннемеловую эпоху наземные формы животных достигли большого разнообразия. На суше по прежнему господствовали хорошо приспособленные к обитанию в лесных и саванных ландшафтах рептилии. Наличие благоприятных условий для жизнедеятельности, обилие пищи и отсутствие врагов способствовали развитию у них гигантизма.
По сравнению с позднеюрской эпохой в раннем мелу число зауропод уменьшилось, господствующее положение заняли птиценогие динозавры - игуанодонты и утконосные динозавры. Господство двуногой динозавровой сухопутной фауны было связано с необходимостью обитания среди высокой растительности и периодическим пребыванием в воде. Эти крупные травоядные животные жили главным образом в болотистых лесах расположенных на берегах крупных пресноводных водоемов Не имея иных средств защиты, они скрывались в озерах, заводях и заливах от хищных динозавров - тарбозавров и тиранозавров. Так же как и позднеюрские предшественники растительноядных динозавров, игуанодонты имели на задних конечностях перепонки, что позволяло им беспрепятственно, несмотря на огромные размеры, передвигаться по влажным и топким грунтам.
Типично сухопутными формами были растительноядные четвероногие рептилии. Для защиты от хищников они имели тяжелые панцири с шипами, роговыми воротниками, колючими (гребнями или толстую шкуру, покрытую мелкими, но плотными роговыми пластинками.
Обилие атмосферных вод и наличие неровностей рельефа способствовали возникновению пресных озер, в которых наряду с различными рыбами обитали довольно многочисленные пресноводные брюхоногие и двустворчатые моллюски. В озерах, зарастающих водной и болотной растительностью (и в связи с этим обедненных кислородом), моллюсковая фауна была угнетенной и весьма малочисленной.
В раннемеловую эпоху среди морской фауны беспозвоночных осуществилась дифференциация по климатическому признаку. В приполярных морях распространялась бореальная фауна, а в низких широтах - тропическая и экваториальная. Четкой границы между этими двумя довольно различными комплексами фауны практически не существовало. В морских бассейнах, занимавших промежуточное положение, обитали смешанные фауны, в составе которых в зависимости от температурных условий преобладали то тропические, то бореальные представители. Бореальная фауна характеризовалась присутствием эндемичных родов аммонитов, белемнитов и двустворчатых моллюсков. Тропические моря были населены богатой фауной беспозвоночных. Основную роль играли кораллы, морские ежи, аммониты, белемниты, брахиоподы, теплолюбивые представители планктонных фораминифер и наннопланктон.
На морском мелководье, там где воды не испытывали сильных изменений солености ввиду отсутствия вблизи речного стока, находились крупные отмели, на которых располагались колонии устриц и рудистов, барьерные и береговые рифы. В районах развития рифовых комплексов обитали брахиоподы, морские ежи, морские лилии и не дожившие до настоящего времени строматопоры. Обширные пространства занимали подводные луга. На некотором удалении от берега находилось основное местообитание белемнитов и аммонитов.
В конце раннего мела ареал обитания бореальной фауны сильно расширился и многие из типичных бореальных форм (например, двустворчатые моллюски рода ауцеллин или аммониты рода краспедитес) достигли тропических морей.
В течение раннемелового времени в результате изменения температурного режима происходило то расширение, то сужение климатических поясов. Существовавший в начале раннемеловой эпохи значительный дефицит влаги постепенно сменился обильным увлажнением в ее конце. Сильно увеличились площади с гумидным типом климата. Одновременно с этим произошло незначительное общее похолодание.
Температуры приповерхностных морских вод в экваториалб ном и тропическом поясах на протяжении большей части позднего мела были довольно высокими. Например, в морях Западной Европы в начале раннего мела они составляли 20-25°С. Примерно такие же температуры существовали в конце раннего мела в морях, расположенных на востоке Южной Африки в Северной Австралии и Северной Америке.
В самом конце раннего мела произошло снижение температур на 2-3°, и это незамедлительно сказалось на составе фауны. Резко сократились ареалы кораллов и рудистов, появились не только организмы, которые могли существовать в различных температурных условиях, но и бореальные организмы, а также уменьшилось карбонатонакопление.
Экваториальный пояс охватывал север Южно-Американского континента, центральную часть Африки, юг Аравии, Индию и Индокитай. По обе стороны от него находились области с тропическим засушливым климатом. Северный аридный сектор охватывал юг США, Мексику, Центральную Америку, север Африки и Южную Евразию, а южный - Южную Америку, Африку и центральные области Австралии. В этих областях преобладали пустыни и полупустыни с ксерофильным редколесьем, весьма сходные с современными саваннами.
Понижение температурного режима в конце раннего мела произошло повсеместно. Так, снижение среднегодовых температур на 5-8°С по сравнению с началом раннемеловой эпохи установлено для морей, расположенных на территории Карпат, Крыма, Северного Кавказа, юга европейской части СССР, Средней Азии и Сибири.
Прохладные условия в течение раннемеловой эпохи существовали в пределах северных окраин Северо-Американского и Евразиатского материков, на юге Африки, Австралии, Южной Америки и в Антарктиде. На севере Евразии среднегодовые температуры в течение продолжительного времени колебались от 12 до 15°С, а в конце раннего мела они составляли 8 - 12°С. Похолодание привело к смещению климатической зональности, и в приполярных районах возник климат, сходный с современным умеренным климатом.
Как уже отмечалось выше, в конце раннего мела произошла существенная перестройка растительного покрова. Этот важнейший в истории Земли и биосферы факт до настоящего времени все еще не нашел полного объяснения. Неясны причины и места возникновения покрытосеменной растительности. По-видимому, в это время произошли достаточно сильные преобразования в природных условиях, которые способствовали широкому распространению покрытосеменной флоры. Последняя была лучше приспособлена к динамическим контрастам ландшафтно-климатических условий, которые можно объяснить изменением состава атмосферы, тектонической активностью планеты и, возможно, космическими причинами. В конце раннего мела продолжались активные перемещения континентов, сильно изменилась концентрация углекислого газа и кислорода в атмосфере, осуществилось глобальное понижение температур, появились сезонные колебания температур и влажности, уменьшилась облачность, а следовательно, усилился приток как яркого солнечного света, так и жесткой радиации. Очевидно, эти факторы и привели к изменению соотношения мало приспособленной к резким колебаниям температур голосеменной и споровой флоры и широкому распространению лучше приспособленной покрытосеменной.
В отличие от голосеменных покрытосеменные растения обладают большей фотосинтезирующей поверхностью, у них более совершенные водно-сосудистая и корневая системы и они могут лучше переносить сезонные колебания температур и влажности. В позднемеловое время хвойно-широколиственные леса произрастали в высоких широтах северного и южного полушарий. В конце позднего мела появились и сильно расширили свои ареалы листопадные теплоумеренные формы - орех, бук и береза. Хвойные были представлены сосновыми (сосна, ель, пихта, тсуга, кедр) и таксодиевыми. Однако еще в течение длительного времени в приполярных районах присутствовали реликты не только раннемеловой, но и даже юрской флоры. Здесь располагались заросли из низкорослых форм цикадофитов, араукариевых и гинкговых. В средних широтах произрастали широколиственные леса - платаны, лавры, дубы, каштаны и клены. В субтропиках росли как широколиственные листопадные, так и вечнозеленые магнолии, лавры, пальмы, а также секвойи, эвкалипты, сосны; в подлеске располагались папоротники, гинкговые и цикадофитовые.
Во влажных тропиках и в приэкваториальной части произрастали дождевые экваториальные леса, вечнозеленая растительность которых была представлена главным образом пальмами, древовидными папоротниками, вечнозелеными дубами, магнолиями и т.д.
В засушливых ландшафтах ксерофильного редколесья и сухих опустыненных саванн господствовали узко- и мелколистные формы растительности. Рощи и небольшие леса встречались лишь в виде оазисов и вдоль речных долин и образовывали галерейные леса.
Среди наземной фауны позвоночных в течение позднего мела появились новые виды четвероногих растительноядных рептилий - анкилозавры , цератопсы , которые обладали пассивными средствами защиты от хищников (шипы, панцирь, роговой покров). Они уже не жили, как их предки, в прибрежно-морских или приозерных зарослях, а переместились в пределы саванного редколесья. В этих ландшафтах водились рогатые цератопсы, очень сходные и в экологическом и в морфологическом отношении с появившимися в кайнозое носорогами. В саванных ландшафтах существовали утконосые гадрозавры. Они перемещались на двух ногах и были тесно связаны с водной средой. Об этом свидетельствуют во-первых, утиная морда с далеко отодвинутыми ноздрями и, во-вторых, наличие плавательных перепонок на ногах Гадрозавры питались прибрежной болотной растительностью обитали в широких речных руслах, дельтах рек и по образу жизни напоминали современных бегемотов.
Большого разнообразия в позднемеловое время достигли крокодилы, основным местообитанием которых являлись реки пойменные озера и старицы. Условия открытой саванны с умеренно аридным климатом и сезонным, но обильным выпадением осадков благоприятствовали развитию динозавровой фауны. Динозавры обитали в многочисленных тихих водоемах, окруженных зарослями из влаголюбивых растений. В заболоченных тропических и субтропических лесах на внутриматериковых аллювиальных и приморских озерно-дельтовых низменностях с обильным увлажнением происходило резкое сокращение видового состава динозавров. В субтропиках это объяснялось не столько высокой влажностью, сколько умеренностью климата и почти сплошным развитием озерно-болотных ландшафтов.
Слабое распространение позвоночных в лесных ландшафтах было связано с неблагоприятными условиями их обитания, непроходимостью лесных зарослей и огромных заболоченных пространств, трудностью пропитания.
В течение позднего мела широкое развитие получили пресноводные моллюски с массивными раковинами. Большого разнообразия достигла фауна морских беспозвоночных, представленная головоногими моллюсками - брахиоподами, двустворчатыми и брюхоногими моллюсками, кораллами, фораминиферами, морскими ежами и лилиями. Распространение всех беспозвоночных определялось температурным режимом. Так, в приполярных районах и морях высоких широт обитал бореальный комплекс, состоящий из песчаных фораминифер и эндемичных видов аммонитов и белемнитов. В нем отсутствовали кораллы и многие моллюски с массивной раковиной. В тропических морях фауна характеризовалась большим разнообразием. Многочисленные кораллы и строматопоры участвовали в построении крупных рифовых массивов. В мелководных морях обитали аммониты, брахиоподы, крупные и мелкие фораминиферы, рудисты и крупные устрицы.
На протяжении позднемеловой эпохи на Земле существовали экваториальный, тропический, субтропический и умеренный пояса. Первые два в отличие от аналогов предшествующей эпохи захватывали более обширные территории. В тропическом поясе обитали теплолюбивые организмы, произрастали вечнозеленые дождевые леса и располагались саванны. Температурный режим повсеместно был довольно высоким. Максимально высокие температуры существовали в середине позднемеловой эпохи. В морях Западной Европы они колебались в пределах 19-25°С, но в конце позднего мела, особенно на границе с кайнозоем, за исключением Южной Европы, среднегодовая температура понизилась до 15°С.
Высокие среднегодовые температуры были характерны для Карпат, Крыма, Закавказья и юга Средней Азии, в течение позднемеловой эпохи они изменялись в пределах 18-27°С. Высокие температуры в течение позднемеловой эпохи существовали в морях, располагавшихся на территории США (20-26°С), на юге Канады (20-28°С) и севере Австралии (20-24°С).
В северном полушарии находились два аридных сектора с тропическим климатом. Один из них охватывал север Африки, Аравию, Восточное Средиземноморье, а другой простирался от Средней Азии через центральные районы Китая до современного побережья Тихого океана. В южном полушарии аридные секторы занимали запад Южной Америки, центральные районы Африки и Индокитай. Для этих областей были характерны пустынные и полупустынные ландшафты и слабое развитие гидрографической сети. В сторону районов с высоким увлажнением пустынные ландшафты сменялись ксерофильным редколесьем.
За пределами тропического пояса располагались районы с относительно низким температурным режимом. На севере европейской части территории СССР, на юге Западной и Восточной Сибири и на Дальнем Востоке среднегодовые температуры изменялись и составляли 14-18°С.
В местах максимального увлажнения, где находились обширные приморские озерно-дельтовые и внутриконтинентальные озерно-аллювиальные низменности и равнины, росли смешанные широколиственные леса с примесью хвойных и папоротников.
Умеренный пояс в северном полушарии охватывал северо-восток Евразии и северо-запад Северной Америки, а в южном полушарии выделялся на юго-востоке Австралии и в Антарктиде. Со временем климат приобрел черты континентальности, и главным показателем этого являются резкие суточные и сезонные перепады температур. Среднегодовые температуры в умеренном поясе обычно составляли 10-14°С, но в отдельные промежутки времени они повышались и это приводило к изменению положения климатических границ.
Гибель гигантов
В конце мелового периода некогда богатая динозавровая фауна пришла в упадок, и ни один из ее представителей не перешел рубеж между мезозойской и кайнозойской эрами. На протяжении сравнительно короткого промежутка времени вымерли не только крупные сухопутные и морские рептилии, но и целый ряд морских беспозвоночных, в частности аммониты, белемниты и некоторые семейства планктонных фораминифер.
Многие поколения ученых питались выяснить причину вымирания динозавров и других представителей мезозойской фауны. Высказывались самые различные предположения - от фантастических до вполне реальных, но и до настоящего времени действительные причины этой катастрофы пока еще не установлены. Ясно лишь то, что причиной гибели не могли послужить какие-либо значительные изменения рельефа земной поверхности, так как он даже в конце мела - начале палеогена оставался равнинным.
В меловом периоде динозавры прочно занимали основные жизненные пространства земной поверхности. И вдруг, а это в геологическом смысле произошло почти мгновенно, как по мановению волшебной палочки, они вымерли. Что же случилось? Конечно, динозавры были примитивнее появившихся им на смену млекопитающих. Они обладали менее энергичным обменом веществ и, следовательно, меньшей активностью, а она во многом зависела от солнечного тепла. Однако эволюционные преобразования и замена более прогрессивными группами в ходе геологической истории должны были происходить постепенно, а не внезапно, как это имело место в конце мезозоя.
Внезапное вымирание динозавров могло быть следствием одновременного наложения многих несомненно взаимосвязанных причин. Наиболее важными могли быть изменения пищевых ресурсов или даже состава пищи, концентрации кислорода и углекислого газа в атмосфере, усиление потока космической радиации и т. д.
Рассмотрим подробнее возможность действия перечисленных причин на жизнедеятельность динозавров. В отличие от сменивших их животных динозавры не имели надежной внешней термоизоляции, как, например, птицы (перья) или млекопитающие (волосяной покров, толстый слой жира). По-видимому, они не имели и постоянной температуры крови, вследствие чего им были опасны как перегрев в жаркое время, так и переохлаждение при значительных понижениях температуры воздуха. Поэтому наиболее распространенным объяснением вымирания динозавров принято считать изменение климатических условий на земной поверхности.
Нормальная жизнедеятельность динозавров протекала в тропическом и экваториальном поясах при равномерной высокой влажности с узкими интервалами в колебаниях суточных, сезонных и годовых температур. Понижение среднегодовых температур до 10-15°С привело к утрате активности и даже способности к передвижению и могло быть причиной гибели динозавров, так как их экологическими преемниками были теплокровные млекопитающие, способные переносить значительные колебания температуры среды обитания. Этот температурный фактор гибели динозавров подтверждается установлением на границе мелового и палеогенового периодов сильного похолодания.
Наибольшие снижения температур примерно до 10-12°С имели место в высоких и средних широтах, в результате чего существовавшие там ландшафты сильно видоизменились. Однако динозавровая фауна вряд ли могла перенести столь значительное похолодание. Тем более что в отличие от млекопитающих динозавры были тесно связаны с местами обитания и не приспособлены к активным миграциям. В тропических областях и приэкваториальной части температуры понизились, но не настолько, чтобы они могли так сильно отразиться на жизнедеятельности крупных организмов. Хорошо известно, что даже во время ледниковых эпох, когда материковое оледенение охватывало значительные территории (например, во время максимального оледенения в четвертичном периоде ледяной панцирь в отдельных районах спускался до 52°с.ш.), в экваториальной части продолжал существовать высокотермический режим, хотя ширина экваториального тропического поясов последовательно суживалась. Следовательно, в конце мелового периода экваториальный и тропический пояса продолжали существовать, и в этих районах должны были сохраниться динозавры. В таком случае могло произойти глобальное вымирание динозавров, а лишь сокращение их ареалов.
Однако динозавры вымерли почти одновременно как в средних, так и в низких широтах. Возможно, определенную роль в гибели рептилий низких широт сыграла наступившая в связи с понижением температур значительная аридизация климата, которая вызвала сильные изменения ландшафтных условий. В конце мелового периода произошла крупная регрессия и обширные участки земной поверхности освободились от водного покрова. Все это привело к резкому увеличению испаряемости, так же губительному для динозавров, как и понижение температур.
Одной из причин вымирания динозавров могло быть измерение интенсивности солнечной радиации и жесткости космических лучей, которое могло произойти на границе мела и палеогена. Некоторые ученые считают, что резкое увеличение интенсивности жесткости космического излучения было вызвано вспышкой сверхновой звезды. Оно оказало губительное влияние на формы жизни, связанные с обитанием на открытых пространствах и в верхней толще воды. В этом случае под воздействием космических лучей должны были погибнуть все живые существа. Однако на суше вымерли только крупные рептилии, а мелкие представители - змеи, ящерицы, черепахи, крокодилы - сохранились. Продолжали существование многие обитатели водной среды, в том числе и жившие в зоне морского мелководья - устрицы, брахиоподы, морские ежи и т. д.
Большой интерес среди геологов вызвала оригинальная гипотеза, согласно которой гибель динозавров произошла вследтвие мутаций в организме, вызванных сильной радиацией: резко возросшая интенсивность космических лучей нарушила воспроизодство потомства. Возникает вопрос: если это в действительности было так, то почему бомбардировка жестким излучением оказалась смертельной только для динозавров? Весьма важное опровержение этой гипотезы было получено в процессе специальных исследований костей ископаемых гигантов. Как оказалось ни кости динозавров, ни их яйца не содержат доказательств повышенной радиоактивности.
Одни ученые усматривали причину гибели динозавров в вирусной эпидемии, а другие считали, что перед гибелью динозавры вдруг стали откладывать яйца с очень толстой скорлупой. В результате этого не все детеныши смогли пробивать скорлупу и рождаемость динозавров резко упала. Однако как в одном, так и а другом случае остается неясным вопрос: почему одновременно вымерли не только наземные, но и водные динозавры?
Как отмечалось выше, динозавры откладывали яйца в теплый песок и под действием определенных температур в них развивались зародыши. Снижение среднегодовых температур и возникновение сезонных и суточных контрастов привело к тому, что яйца не получали необходимого тепла и зародыши в них в конце концов погибали. Поэтому рождаемость динозавров резко снизилась, и в дальнейшем они вымерли.
Недавно была высказана гипотеза, суть которой заключается в том, что динозавры погибли в результате массового отравления алкалоидами покрытосеменных растений. Однако этому предположению противоречит тот факт, что массовое появление покрытосеменных приходится на конец раннемеловой эпохи, а гибель динозавров произошла только в конце позднемеловой эпохи, т. е. примерно через 30 - 40 млн. лет. Если алкалоиды покрытосеменных были действительно смертельными для динозавров, то они должны были исчезнуть уже в начале позднего мела, однако они продолжали еще долго жить и развиваться. Так что и эта гипотеза не в состоянии объяснить причину массового вымирания динозавров.
Существует еще одна гипотеза, с помощью которой пытаются объяснить гибель динозавров в результате изменения содержания химических соединений и микроэлементов в окружающей среде. Появление новых микроэлементов и соединений могло сильно повлиять на судьбу динозавров. Многие элементы и соединения освобождались при разложении горных пород под воздействием на них природных вод, обогащенных органическими кислотами и углекислым газом. Легкорастворимые вещества при выветривании выносились, а соединения железа (преимущественно гидроокислы железа), алюминия и титана оставались на месте. Известно, что карбонат кальция и фосфаты являются необходимой составной частью скелетных образований животных. Наибольший недостаток этих соединений ощущается, в тропическом и экваториальном поясах, т. е. в главном местообитании динозавров. Животный мир современных тропических влажных лесов характеризуется небольшими размерами, что является своеобразной реакцией органического мира на дефицит карбоната кальция и фосфатов. Продолжающееся в течение всей позднемеловой эпохи интенсивное выветривание привело к тому, что почвенные воды и растительность оказались обеднены этими соединениями. Динозавры стали испытывать сильный известковый и фосфатный голод, кости их, не получая в достаточном количестве эти соединения, не могли выдерживать больших нагрузок, что и привело к массовой гибели гигантов.
В процессе интенсивного выветривания нарушился круговорот микроэлементов, играющих большую роль в жизнедеятельности животных. Уменьшение количества натрия в крови вызывало истощение нервной системы, дефицит железа способствовал развитию малокровия, уменьшение йода влекло за собой увеличение щитовидной железы, а нехватка фтора способствовала развитию болезней зубов. Поэтому большой дефицит микроэлементов мог вызвать массовую гибель таких животных, как гигантские ящеры. Сильное влияние на развитие животных оказывает присутствие в почвенных растворах, корах выветривания и морских бассейнах стронция. Повышение его концентрации приводит к тому, что он замещает кальций в костной ткани. В то же время стронций довольно быстро удаляется из тканей и костей животных, и кости становятся пористыми и хрупкими.
Не менее вредное воздействие на организм животных оказывают и другие элементы. Переход селена из почв и растений в организмы животных приводит к нарушению работы сердца, поражению суставов, выпадению волосяного покрова и т. д. Недостаток кобальта в организме животных вызывает авитаминоз, а меди - малокровие.
Вероятно, в конце мезозоя в силу взаимного наложения многих причин произошла гибель гигантов. Вполне возможно, что главной причиной вымирания динозавров было изменение климата, сопровождаемое последовательными изменениями других природных условий.
В процессе глубоководного бурения в океанах на границе между меловыми и палеогеновыми отложениями был обнаружен тонкий слой глины, обогащенный иридием. В связи с этим ученые высказали предположение, что динозавры могли в результате отравления иридием и другими тяжелыми металлами. Повышение содержания иридия в породах объясняется столкновением нашей планеты с крупным космическим телом. Недавно слой, содержащий повышенное количество иридия на границе мела и палеогена, обнаружен в Италии, Югославии и СССР.
| <<Назад |
Мезозойская эра,продолжительность которой составляет 183 млн лет, подразделяется на три периода - триасовый, юрский и меловой. Соответственно подразделяется на системы и мезозойская группа отложений.
Т р и а с о в а я система получила свое название в связи с четким подразделением ее отложений на три части - нижний, средний и верхний триас. Соответственно, триасовый период (35,0 млн. лет) делится на три отдела - ранний, средний и поздний.
В мезозое континенты Северного и Южного полушарий разделялись вытянутым в широтном направлении обширным морским бассейном. Он получил название Тетис - в честь древнегреческой богини моря.
В начале триаса в некоторых районах земного шара происходили мощные вулканические извержения. Так, в Восточной Сибири излияния базальтовой магмы образовали толщу основных горных пород, залегающих в виде огромных покровов. Такие покровы называются "траппами " (швед. "траппа" - лестница ). Для них характерна столбчатая отдельность в виде ступеней лестницы. Вулканические извержения происходили также в Мексике и на Аляске, в Испании и Северной Африке. В Южном полушарии триасовый вулканизм резко проявился в Новой Каледонии, Новой Зеландии, Андах и других районах.
В триасе произошла одна из крупнейших в истории Земли регрессий моря. Она совпала с началом новой складчатости, продолжавшейся в течение всего мезозоя и получившей название "мезозойской". Возникшие в это время складчатые сооружения получили название "мезозоид".
Ю р с к а я система названа по Юрским горам в Швейцарии. В юрский период, продолжавшийся 69,0 млн. лет, началась новая трансгрессия моря. Но в конце юры в области океана Тетис (Крым, Кавказ, Гималаи и др.) и особенно ощутимо в области Тихоокеанских окраин возобновились горообразовательные движения. Они привели к образованию горных сооружений внешнего Тихоокеанского кольца: Верхоянско-Колымских, Дальневосточных, Андийских, Кордильерских. Складчатость сопровождалась активной вулканической деятельностью. В Южной Африке и Южной Америке (бассейн реки Парана) в начале юрского периода произошли крупные излияния основных лав траппового характера. Мощность базальтовых толщ здесь достигает более 1000 метров.
М е л о в а я система получила свое название в связи с тем, что в ее отложениях широко распространены слои белого мела. Меловой период продолжался 79,0 млн. лет. Его начало совпало с обширнейшей морской трансгрессией. Согласно одной из гипотез, северный сверхматерик Лавразия в это время распался на ряд отдельных континентов: Восточно-Азиатский, Северо-Европейский, Северо-Американский. Гондвана также распалась на отдельные континентальные массивы: Южно-Американский, Африканский, Индостанский, Австралийский и Антарктический. В мезозое образовались, возможно, все современные океаны, кроме, видимо, более древнего Тихого океана.
В позднемеловую эпоху на территориях, прилегающих к акватории Тихого океана, проявилась мощная фаза мезозойской складчатости. Менее интенсивные горообразовательные движения в это время происходили в ряде районов Средиземноморской области (Восточные Альпы, Карпаты, Закавказье). Как и в юрский период, складчатость сопровождалась интенсивным магматизмом.
Мезозойские породы "пронзены" внедришимися в них гранитными интрузиями. А на обширных пространствах Сибирской, Индийской, Африкано-Аравийской платформ в конце мезозоя происходили грандиозные излияния базальтовых лав, сформировавших трапповые покровы (швед. «трапп» - лестница ). Ныне они выходят на поверхность, например, по берегам реки Нижней Тунгуски. Здесь можно наблюдать возвышающиеся на несколько сотен метров останцы твердых базальтов, которые были внерены ранее в осадочные породы, уничтоженные после выхода на поверхность процессами выветривания и размыва. Вертикальные уступы черных (темно-серых), называемых «столбами», траппов чередуются с горизонтальными площадками. Этим они полюбились альпинистам и туристам. Мощность таких покровов на Деканском плато в Индостане достигает 2000-3000 м.
О р г а н и ч и ч е с к и й м и р м е з о з о я. На рубеже палеозойской и мезозойской эр существенно обновилсяживотный и растительный мир (рис. 14, 15). Для триасового периода характерно появление в морях новых головоногих (аммонниты, белемниты) и пластинчатожаберных моллюсков, шестилучевых кораллов и других групп животных. Появились костистые рыбы.
На суше это было время господства пресмыкающихся. Возникли новые их группы - первые ящерицы, черепахи, крокодилы, змеи. В начале мезозоя появились и первые млекопитающие - мелкие сумчатые величиной в современную крысу.
В триасе - юре появились и расцвели белемниты, гигантские растительноядные и хищные пресмыкающиеся ящеры - динозавры (греч. "динос" - страшный, "саврос" - ящер). Они достигали в длину 30 м и более и весили до 60 тонн. Динозавры (рис. 16) освоили не только сушу, но и море. Здесь обитали ихтиозавры (греч. "ихтис" - рыба) - крупные хищные рыбоящеры, достигавшие в длину более 10 м и походившие на современных дельфинов. Тогда же появились первые летающие ящеры - птерозавры (греч. "птерон" - крыло) , "саврос" - ящер). Это были в основном небольшие (до полуметра) пресмыкающиеся, приспособившиеся к полету.
Распространенными представителями птерозавров являлись летающие ящеры - рамфоринхи (греч. рамфос" - клюв, "ринос" - нос) и птеродактили (греч. "птерон" - перо, "дактилос" - палец). Их передние конечности превратились в летательные органы - перепончатые крылья. Главную пищу рамфоринхов составляли рыбы и насекомые. Наиболее мелкие птеродактили были величиной с воробья, наиболее крупные достигали размеров ястреба.
Летающие ящеры не являлись предками птиц. Они представляют собой особую, самостоятельную эволюционную ветвь пресмыкающихся, которая полностью вымерла в конце мелового периода. Птицы произошли от других пресмыкающихся.
Самой первой птицей, видимо, является археоптерикс (греч. "археос" - древний, "птерон" - крыло). Это была переходная форма от пресмыкающихся к птицам. Размером археоптерикс был с ворону. Он имел короткие крылья, острые хищные зубы и длинный хвост с веерообразным оперением. Формой тела, строением конечностей и наличием оперения археоптерикс был сходен с птицами. Но по ряду признаков был еще близок к пресмыкащимся.
В юрских отложениях обнаружены остатки примитивных млекопитающих.
Меловой период - время наибольшего расцвета пресмыкающихся. Динозавры достигли огромных размеров (до 30 м в длину); масса их превышала 50 т. Они широко заселили сушу и воды, царили в воздухе. Летающие ящеры в меловой период достигли гигантских размеров - с размахом крыльев около 8 м.
Гигантские размеры были свойственны в мезозое и некоторым другим группам животных. Так, в меловых морях существовали моллюски - аммониты, раковины которых достигали в диаметре 3 м.
Из растений на суше, начиная с триасового периода, преобладали голосеменные: хвойные, гингковые и др.; из споровых - папоротники. В юрский период бурное развитие получила наземная растительность. В конце мелового периода появились покрытосеменные растения; на суше образовался травяной покров.
В конце мелового периода органический мир снова претерпел резкие изменения. Вымерли многие беспозвоночные и большинство гигантских ящеров. Причины их вымирания достоверно не установлены. Согласно одной гипотезе, гибель динозавров связывают с геологической катастрофой, происшедшей около 65 млн. лет назад. Предполагают, что тогда с Землей столкнулся крупный метеорит.
В 70-х годах ХХ в. геолог Калифорнийского университета Уолтер Альварес и
его отец физик Луис Альварес обнаружили в пограничных мел-палеогеновых отложениях разреза Губбио (Италия) необычайно высокое содержание иридия - элемента, в большом количестве содержащегося в метеоритах. Аномальное содержание иридия было обнаружено также на границе мела и палеогена в других
районах земного шара. В связи с этим отец и сын Альваресы выдвинули гипотезу о столкновении с Землей крупного космического тела астероидного размера. Следствием столкновения было массовое вымирание мезозойских растений и животных, в частности динозавров. Это произошло около 65 млн. лет назад на рубеже мезозойской и кайнозойской эр. В момент столкновения мириады метеоритных частиц и земного вещества поднялись гигантским облаком в небо и на годы закрыли Солнце. Земля погрузилась в темноту и холод.
В первой половине 80-х годов были проведены многочисленные геохимические исследования. Они показали, что содержание иридия в пограничных мел-палеогеновых отложениях действительно очень высоко - на два-три порядка выше его среднего содержания (кларка) в земной коре.
В конце позднего исчезли и большие группы высших растений.
П о л е з н ы е и с к о п а е м ы е м е з о з о я.
Отложения мезозоя содержат много полезных ископаемых. Месторождения рудных полезных ископаемых образовывались в результате проявления базальтового магматизма.
В широко распространенной триасовой коре выветривания присутствуют месторождения каолинов и бокситов (Урал, Казахстан). В юрский и меловой периоды происходило мощное угленакопления. В России месторождения мезозойских бурых углей расположены в пределах Ленского, Южно-Якутского, Канско-Ачинского, Черемховского, Чулымо-Енисейского, Челябинского бассейнов, на Дальнем Востоке и в других районах.
К юрским и меловым отложениям приурочены знаменитые месторождения нефти и газа Ближнего Востока, Западной Сибири, а также Мангышлака, Восточной Туркмении и Западного Узбекистана.
В юрский период образовались горючие сланцы (Поволжье и Общий Сырт), осадочные железные руды (Тульская и Липецкая области), фосфориты (Чувашия, Подмосковье, Общий Сырт, Кировская область).
К меловым отложениям приурочены месторождения фосфоритов (Курская, Брянская, Калужская и др. области) и бокситов (Венгрия, Югославия, Италия, Франция). С меловыми гранитными интрузиями и базальтовыми излияниями связаны месторождения полиметаллических руд (золота, серебра, меди, свинца, цинка, олова, молибдена, вольфрама и др.). Это, например, Садонское (Северный Кавказ) месторождение полиметаллических руд, оловянные руды Боливии и т.п. По берегам Тихого океана простираются два богатейших мезозойских рудных пояса: от Чукотки до Индокитая и от Аляски до Центральной Америки. В Южной Африке и Восточной Сибири к меловым отложениям приурочены месторождения алмазов.
Кайнозойская эра. Кайнозойская эра продолжается 65 млн. лет. В международной шкале геологического времени она подразделяется на "третичный" и "четвертичный" периоды. В России и других государствах бывшего Советского Союза кайнозой подразделяется на три периода: палеогеновый, неогеновый и антропогеновый (четвертичный).
П а л е о г е н о в ы й период (40,4 млн. лет) делится на раннюю - палеоценовую (10,1 млн. лет), среднюю - эоценовую (16,9 млн. лет) и позднюю - олигоценовую (13,4 млн. лет) эпохи. В Северном полушарии в палеогене существовали Северо-Американский и Евразиатский материки. Их разделяла впадина Атлантического океана. В Южном полушарии продолжали развиваться самостоятельно материки, отколовшиеся от Гондваны и разделенные впадинами Атлантического и Индийского океанов.
В эоценовую эпоху в области Средиземноморья проявилась первая фаза мощной альпийской складчатости. Она вызвала поднятие некоторых центральных участков этой области. К концу палеогена море полностью покинуло территорию Гималайско-Индостанской части Тетиса.
Образование многочисленных глубинных разломов в области Северного пролива и прилегающих районов Ирландии, Шотландии, Северной Англии и Гебридских островов; района Южной Швеции и Скагеррака, а также во всей области Северной Атлантики (Шпицберген, Исландия, Западная Гренландия) способствовало базальтовым излияниям.
В конце палеогенового периода во многих частях земного шара широко проявились разрывные и блоковые движения земной коры. В ряде районов Западно-Европейских герцинид возникла система грабенов (Верхнерейнский, Нижнерейнский). Система узких меридионально вытянутых грабенов (Мертвое и Красное моря, озера Альберта, Ньяса, Танганьика) возникла в восточной части Африканской платформы). Она протянулась от северной окраины платформы почти до крайнего юга на расстоянии свыше 5000 км. Разрывные дислокации здесь сопровождались грандиозными излияниями базальтовых магм.
Н е о г е н о в ы й период включает две эпохи: раннюю - миоценовую (19,5 млн. лет) и позднюю - плиоценовую (3,5 млн. лет). Для неогена было характерно активное горообразование. К концу неогена альпийская складчатость превратила большую часть области Тетиса в наиболее молодую в структуре земной коры альпийскую складчатую область. В это время приобрели свой современный облик многие горные сооружения. Возникли цепи Зондских, Молуккских, Ново-Гвинейских, Ново-Зеландских, Филиппинских, Рюккю, Японских, Курильских, Алеутских островов и др. В пределах Восточно-Тихоокеанских прибрежных окраин узкой полосой поднялись береговые хребты. Горообразование происходило и в области Центрально-Азиатского горного пояса.
Мощные блоковые движения вызвали в неогене опускание крупных участков земной коры - областей Средиземноморского, Адриатического, Черного, Восточно-Китайского, Южно-Китайского, Японского, Охотского и других краевых морей, а также Каспийского моря.
Поднятия и опускания блоков земной коры в неогене сопровождались
зарождением глубинных разломов. По ним происходило излияние лав. Например,
в области Центрального плато Франции. В зоне этих разломов возникли в неогене вулканы Везувий, Этна, а также камчатские, курильские, японские и яванские вулканы.
В истории Земли были часты периоды похолодания, чередовавшиеся с потеплением. Около 25 млн. лет назад, с конца палеогена, произошло похолодание. Одно из потеплений имело место в начале позднего неогена (плиоценовая эпоха). Следующее похолодание сформировало горно-долинные и покровные ледники в северном полушарии и мощный ледяной покров в Арктике. Многолетнее промерзание пород на севере России продолжается до настоящего времени.
А н т р о п о г е н о в ы й период получил свое название потому, что в начале этого периода появился человек (греч. "антропос" - человек ). Его прежнее название - четвертичная система. Вопрос о длительности антропогенового периода до настоящего времени окончательно не решен. Одни геологи определяют длительность антропогена не менее 2 млн лет. Антропоген подразделяется на эоплейстоцен (греч. "эос" - заря, "плейстос" - наибольший, "кайнос" - новый ), плейстоцен и голоцен (греч. "голос" - весь, "кайнос" - новый ). Длительность голоцена не превышает 10 тыс. лет. Но некоторые ученые относят эоплейстоцен к неогену и нижнюю границу антропогена проводят на уровне 750 тыс. лет назад.
В это время более активно продолжилось поднятие Центрально-Азиатского горно-складчатого пояса. По мнению некоторых ученых, горы Тянь-Шаня и Алтая за антропогеновый период поднялись на несколько километров. А впадина озера Байкал погрузилась на 1600 м.
В антропогене проявляется интенсивная вулканическая деятельность. Наиболее мощные базальтовые излияния в современную эпоху наблюдаются в средино-океанических хребтах и других обширных пространствах океанского дна.
"Великие" оледенения происходили на огромных пространствах северных материков и в антропогеновый период. Они сформировали и ледниковый покров Антрактиды. Эоплейстоцен и плейстоцен характеризуются общим похолоданием климата Земли и периодическим возникновением материковых оледенений в средних широтах. В среднем плейстоцене мощные ледниковые языки спустились почти до 50° с.ш. в Европе и до 40° с.ш. в США. Здесь мощность моренных отложений составляет первые десятки метров. Межледниковые эпохи характеризовались относительно мягким климатом. Средние температуры повышались на 6 - 12° С (Н.В.Короновский, А.Ф.Якушова, 1991). .
Формировавшиеся за счет вод морей и океанов огромные массы льда в виде ледников надвигались на сушу. Мерзлые породы распространялись на обширные пространства. Голоцен - послеледниковая эпоха. Его начало совпадает с окончанием последнего материкового оледенения Северной Европы.
О р г а н и ч и ч е с к и й м и р к а й н о з о я. К началу кайнозойской эры вымирают белемниты, аммониты, гигантские пресмыкающиеся и др. В кайнозое активно стали развиваться простейшие (фораминиферы), млекопитающие и костистые рыбы. Они заняли господствующее положение среди других представителей животного мира. В палеогене среди них преобладали яйцекладущие и сумчатые (подобие фауны этого типа частично сохранилось в Австралии). В неогене эти группы животных отступают на второй план и основную роль начинают играть копытные, хоботные, хищники, грызуны и другие известные ныне классы высших млекопитающих.
Органический мир антропогена похож на современный. В антропогеновый период от приматов, существовавших еще в неогене 20 млн. лет назад, произошел человек.
Кайнозойская эра характеризуется широким распространением наземной растительности: покрытосемянных растений, трав, близких к современным.
П о л е з н ы е и с к о п а е м ы е к а й н о з о я. В палеогеновый период происходило мощное углеобразование. Месторождения бурых углей известны в палеогене Кавказа, Камчатки, Сахалина, США, Южной Америки, Африки, Индии, Индокитая, Суматры. Палеогновые марганцевые руды выявлены на Украине (Никополь), в Грузии (Чиатура), на Северном Кавказе, Мангышлаке. Известны палеогеновые месторождения бокситов (Чулымо-Енисейское, Акмолинское), нефти и газа.
К неогеновым отложениям приурочены залежи нефти и газа (Баку, Майкоп, Грозный, Юго-Западная Туркмения, Западная Украина, Сахалин). В бассейне Черного моря, на территории Керченского и Таманского полуострова, в неогеновый период в различных районах происходило осаждение железных руд.
В антропогеновый период образовались месторождения солей, строительных материалов (щебень, гравий, песок, глина, суглинок), озерно-болотных железных руд; а также россыпные месторождения золота, платины, алмазов, оловянной, вольфрамовой руд, драгоценных камней и др.

Меловой период, или мел (145-66 млн лет назад)
Последний период мезозоя назывался мелом. В произошел рост цветущих наземных растений. Им помогали недавно появившиеся пчелы и теплые климатические условия. Хвойные растения все еще были многочисленными в течение мела.
Что касается морских животных мелового периода, акулы и скаты стали обычным явлением. , пережившие пермское вымирание, такие как морские звезды, также были в обилии в течение мела.
На суше первые мелкие млекопитающие начали развиваться в меловой период. Сначала появились сумчатые, а затем и другие млекопитающие. Появилось больше птиц и стало больше рептилий. Продолжалось доминирование динозавров, увеличилось число плотоядных видов.
В конце мела и мезозоя произошло еще одно . Это исчезновение обычно называется K-T вымиранием (мел-палеогеновое вымирание). Оно уничтожило всех динозавров, кроме птиц, и многих других форм жизни на Земле.
Существуют разные версии относительно того, почему произошло массовое исчезновение. Большинство ученых согласны, что это было какое-то катастрофическое событие, вызвавшее это вымирание. Различные гипотезы включают в себя массивные извержения вулканов, из-за которых в атмосферу попало огромное количество пыли, что уменьшило количество солнечного света, достигающего поверхности Земли и тем самым вызвало гибель фотосинтезирующих организмов, таких как растения, и тех, кто от них зависел. Другие считают, что на Землю упал метеорит, а пыль закрыла солнечный свет. Поскольку растения и животные, которые ими питались вымерли, это привело к тому, что хищники, такие как плотоядные динозавры, также погибли из-за отсутствия пищи.
Мезозойская эра делится на триасовый, юрский и меловой периоды общей продолжительностью 173 млн. лет. Отложения этих периодов составляют соответствующие системы, которые в совокупности образуют мезозойскую группу. Триасовая система выделена в Германии, юрская и меловая - в Швейцарии и Франции. Триасовая и юрская системы делятся на три отдела, меловая - на два.
Органический мир
Органический мир мезозойской эры сильно отличается от палеозойской. На смену вымершим в перми палеозойским группам появились новые - мезозойские.
В мезозойских морях получили исключительное развитие головоногие моллюски - аммониты и белемниты, резко увеличилось разнообразие и количество двустворчатых и брюхоногих моллюсков, появились и развивались шестилучевые кораллы. Из позвоночных широко распространились костистые рыбы и плавающие рептилии.
На суше господствовали чрезвычайно разнообразные пресмыкающиеся (особенно динозавры). Среди наземных растений испытали расцвет голосеменные.
Органический мир триасового периода. Особенностью органического мира этого периода являлось существование некоторых архаических палеозойских групп, хотя преобладали уже новые - мезозойские.
Органический мир моря. Среди беспозвоночных были широко распространены головоногие и двустворчатые моллюски. Среди головоногих господствовали цератиты, которые вытеснили гониатитов. Характерным родом был цератитес с типичной цератитовой перегородочной линией. Появились первые белемниты, но в триасе их было еще мало.
Двустворчатые моллюски заселили богатые пищей мелководные участки, на которых в палеозое обитали брахиоподы. Двустворки быстро развивались, становились более разнообразны по составу. Увеличилось число брюхоногих моллюсков, появились шестилучевые кораллы и новые морские ежи с прочным панцирем.
Продолжали развиваться морские позвоночные. Среди рыб сократилось число хрящевых, стали редки кистеперые и двоякодышащие. На смену им появились костистые рыбы. В морях обитали первые черепахи, крокодилы и ихтиозавры - крупные плавающие ящеры, похожие на дельфинов.
Органический мир суши тоже изменился. Вымирали стегоцефалы, а пресмыкающиеся становились господствующей группой. На смену вымирающим котилозаврам и зверообразным ящерам пришли мезозойские динозавры, особенно широко распространившиеся в юре и мелу. В конце триаса появились первые млекопитающие, они имели небольшие размеры и примитивное строение.
Растительный мир в начале триаса был сильно обеднен, сказывалось влияние засушливого климата. Во второй половине триаса климат увлажнился, появились разнообразные мезозойские папоротники и голосеменные (цикадовые, гинкговые и др.). Наряду с ними были широко распространены хвойные. К концу триаса флора приобрела мезозойский облик, характеризующийся господством голосеменных.
Органический мир юрского периода
Органический мир юры был наиболее типичным для мезозойской эры.
Органический мир моря. Среди беспозвоночных господствовали аммониты, они имели сложную перегородочную линию и были чрезвычайно разнообразны по форме раковины и ее скульптуре. Одним из типичных позднеюрских аммонитов является род виргатитес со свойственными только ему пучками ребер на раковине. Много стало белемнитов, их ростры встречаются в массовом количестве в юрских глинах. Характерными родами являются цилиндротеутис с длинным цилиндрическим ростром и гиболитес с ростром веретенообразной формы.
Двустворчатые и брюхоногие моллюски стали многочисленны и разнообразны. Среди двустворок было много устриц с толстой раковиной разнообразной формы. В морях обитали различные шестилучевые кораллы, морские ежи и многочисленные простейшие.
Среди морских позвоночных продолжали господствовать рыбоящеры - ихтиозавры, появились чешуйчатые ящеры - мезозавры, похожие на гигантских зубастых ящериц. Быстро развивались костистые рыбы.
Органический мир суши был очень своеобразным. Безраздельно господствовали гигантские ящеры - динозавры - разнообразной формы и размеров. С первого взгляда они кажутся пришельцами из внеземного мира или плодом фантазии художников.
Наиболее богаты остатками динозавров пустыня Гоби и соседние участки Центральной Азии. Эта огромная территория в течение 150 млн. лет до юрского периода находилась в континентальных условиях, благоприятных для длительного развития ископаемой фауны. Предполагают, что эта область явилась очагом возникновения динозавров, откуда они расселились по всему свету вплоть до Австралии, Африки, Америки.
Динозавры имели гигантские размеры. Современные нам слоны - самые крупные сегодня из наземных животных (ростом до 3,5 м и массой до 4,5 т) - кажутся карликами по сравнению с динозаврами. Наиболее крупными были растительноядные динозавры. «Живые горы» - брахиозавры, бронтозавры и диплодоки - имели длину до 30 м и достигали 40-50 т. Огромные стегозавры несли на спине большие (до 1 м) костные пластины, которые защищали их массивное тело. На конце хвоста стегозавров находились острые шипы. Среди динозавров было немало страшных хищников, которые передвигались гораздо быстрее своих травоядных сородичей. Размножались динозавры при помощи яиц, зарывая их в горячий песок, как это делают современные черепахи. В Монголии до сих пор находят древние кладки яиц динозавров.
Воздушную среду освоили летающие ящеры - птерозавры с острыми перепончатыми крыльями. Среди них выделялись рамфоринхи - зубастые ящеры, питавшиеся рыбой и насекомыми. В конце юры появились первоптицы - археоптериксы - величиной с галку, они сохранили многие черты своих предков - пресмыкающихся.
Растительный мир суши отличался расцветом разнообразных голосеменных: цикадовых, гинкговых, хвойных и др. Юрская флора была довольно однородной на земном шаре и только в конце юры начали намечаться флористические провинции.
Органический мир мелового периода
В течение этого периода органический мир претерпел существенные изменения. В начале периода он был сходен с юрским, а в позднем мелу стал резко сокращаться за счет вымирания многих мезозойских групп животных и растений.
Органический мир моря . Среди беспозвоночных были распространены те же группы организмов, как и в юрском периоде, но состав их изменился.
Продолжали господствовать аммониты, среди них появилось много форм с частично или почти полностью развернутой раковиной. Известны меловые аммониты со спирально-конической (как улитки) и палкообразной раковиной. В конце периода все аммониты вымерли.
Белемниты достигли своего расцвета, они были многочисленны и разнообразны. Особенно широко был распространен род белемнителла с ростром, похожим на сигару. Увеличивалось значение двустворчатых и брюхоногих моллюсков, они постепенно захватывали господствующее положение. Среди двустворок было много устриц, иноцерамусов и пектенов. В тропических морях позднего мела обитали своеобразные бокаловидные гиппуриты. По форме своей раковины они напоминают губок и одиночных кораллов. Это является доказательством того, что эти двустворчатые моллюски вели прикрепленный образ жизни, в отличие от своих сородичей. Большого разнообразия достигли брюхоногие моллюски, особенно к концу периода. Среди морских ежей господствовали различные неправильные ежи, одним из представителей которых является род микрастер с раковиной сердцевидной формы.
Тепловодные позднемеловые моря были переполнены микрофауной, среди которой преобладали мелкие фораминиферы-глобигерины и ультрамикроскопические одноклеточные известковые водоросли - кокколитофориды. Скопление кокколитов образовывало тонкий известковый ил, из которого в дальнейшем формировался писчий мел. Наиболее мягкие разности писчего мела почти целиком состоят из кокколитов, примесь фораминифер в них незначительна.
В морях было много позвоночных. Быстро развивались костистые рыбы, и они завоевывали морскую среду. До конца периода существовали плавающие ящеры - ихтиозавры, мозозавры.
Органический мир суши в раннем мелу мало отличался от юрского. В воздухе господствовали летающие ящеры - птеродактили, похожие на гигантских летучих мышей. Размах их крыльев достигал 7-8 м, а в США был обнаружен скелет гигантского птеродактиля с размахом крыльев 16 м. Наряду с такими громадными летающими ящерами обитали птеродактили размером не более воробья. На суше продолжали господствовать различные динозавры, но в конце мелового периода все они вымерли вместе со своими морскими сородичами.
Наземная флора раннемеловой эпохи, как и в юре, характеризовалась господством голосеменных растений, но начиная с конца раннего мела появляются и быстро развиваются покрытосеменные, которые вместе с хвойными становятся господствующей группой растений к концу мелового периода. Голосеменные резко сокращаются в количестве и разнообразии, многие из них вымирают.
Таким образом, в конце мезозойской эры произошли существенные изменения как в животном, так и в растительном мире. Исчезли все аммониты, большинство белемнитов и брахиопод, все динозавры, крылатые ящеры, многие водные рептилии, древние птицы, ряд групп высших растений из голосеменных.
Среди этих существенных изменений особенно поражает быстрое исчезновение с лица Земли мезозойских гигантов - динозавров. Что же явилось причиной гибели столь большой и разнообразной группы животных? Эта тема давно привлекает ученых и до сих пор не сходит со страниц книг и научных журналов. Существует несколько десятков гипотез, и появляются новые. В основу одной группы гипотез положены тектонические причины - сильный орогенез вызвал существенные изменения палеогеографии, климата и пищевых ресурсов. Другие гипотезы связывают гибель динозавров с процессами, происходившими в космосе, главным образом с изменением космической радиации. Третья группа гипотез объясняет гибель гигантов различными биологическими причинами: несоответствие между объемом мозга и весом тела животных; быстрым развитием хищных млекопитающих, поедавших мелких динозавров и яйца крупных; постепенным утолщением скорлупы яиц до такой степени, что детеныши не могли ее пробить. Существуют гипотезы, связывающие гибель динозавров с увеличением микроэлементов в окружающей среде, с кислородным голоданием, с вымыванием извести из почвы или с возрастанием силы тяжести на Земле до такой степени, что гиганты-динозавры были раздавлены собственным весом.
Трудно отдать предпочтение какой-либо из этих гипотез. По-видимому, в каждой из них есть большая или меньшая доля правды (за исключением совершенно невероятных предположений, которые также высказывались). Несомненно, целый комплекс причин вызвал гибель динозавров, а также сильные изменения в составе других групп животного и растительного мира.
Структура земной коры и палеогеография в начале эры
В начале мезозойской эры структура земной коры сильно отличалась от структуры, существовавшей в начале позднего палеозоя. Большие изменения произошли в строении геосинклинальных поясов. В результате герцинской складчатости завершилось геосинклинальное развитие Урало-Монгольского, Атлантического и Арктического поясов, частей Тихоокеанского и Средиземноморского поясов. С начала мезозоя все эти колоссальные по площади территории превратились в молодые платформы, которые соединили древние платформы северного полушария в огромный платформенный массив Лавразия, отделявшийся от Гондваны геосинклинальными областями Средиземноморского пояса. Геосинклинальный режим продолжал существовать только в Средиземноморском и Тихоокеанском поясах на значительно меньших площадях, чем в начале палеозойской эры, так как отдельные области этих поясов превратились в молодые платформы.
Все эти изменения в структуре земной коры сильно отразились на палеогеографии. В начале мезозойской эры оба платформенных массива - Лавразия и Гондвана находились в континентальных условиях, моря располагались в геосинклинальных областях и заливали краевые части платформ.
Существовала впадина Тихого океана и начала формироваться впадина Индийского океана.
История геологического развития геосинклинальных поясов
Геосинклинальные пояса в течение мезозоя развивались по-разному. На значительных территориях Средиземноморского и Тихоокеанского поясов продолжалось геосинклинальное развитие. Существенным различием в истории этих поясов было интенсивное проявление мезозойской складчатости в Тихоокеанском, в то время как в Средиземноморском поясе она сказалась гораздо слабее и охватила меньшие площади.
Все другие пояса в течение мезозоя развивались как молодые платформы.
Средиземноморский геосинклинальный пояс
Геосинклинальные условия в течение всей мезозойской эры сохранялись на обширной территории пояса в пределах двух геосинклинальных областей - Альпийско-Гималайской и Индонезийской. Альпийско-Гималайская область охватывала западную и центральную части пояса (Южная Европа, Северо-Западная Африка, Малая Азия, Гималаи); Индонезийская - восточную (Бирма, Индонезия, часть Филиппинских островов). Мезозойская складчатость проявилась широко на территории пояса, но складчатые структуры - мезозоиды - возникли только на востоке (Южный Тибет, Индокитай, Малакка).
Краевые части Средиземноморского пояса, которые в результате герцинской складчатости превратились в складчатые структуры - герциниды (значительные территории Западной Европы, Северо-Западной Африки и т. д.), развивались в мезозое как молодые платформы.
Альпийско-Гималайская геосинклинальная область. Мезозойская история этой огромной области исключительно сложна. Она хорошо восстановлена в южной части Западной Европы и на Кавказе. Для понимания основных закономерностей развития Альпийско-Гималайской области в мезозое кратко рассмотрим историю развития Кавказа, изученную с большой полнотой.
Начиная с мезозоя на Кавказе существовали две геосинклинальные системы. На севере располагалась система Большого Кавказа, она охватывала территорию современного Большого Кавказского хребта и протягивалась в пределы Горного Крыма. На юге - система Малого Кавказа, в состав которой входили Армения, южные части Азербайджана и Грузии. Геологическая история этих геосинклинальных систем существенно различалась.
Триасовая история Кавказавосстановлена плохо, так как триасовые отложения встречены на весьма ограниченных участках. Породы представлены преимущественно известняками. По-видимому, на Кавказе в триасе существовал мелководный морской бассейн, но значительные части территории представляли собой сушу. Морской бассейн в течение триаса постепенно расширялся.
Юрская и меловая история Кавказа хорошо восстановлена, она протекала в сложных условиях. Именно в эти периоды формировались и развивались обе геосинклинальные системы. В ранней юре на месте Большого Кавказа существовал широкий и глубокий прогиб, в котором накапливалась очень мощная толща глин с прослоями песчаников. В средней юре этот прогиб разделился на две части, в его центре возникло геоантиклинальное поднятие. В северном прогибе продолжалось накопление очень мощных песчано-глинистых осадков (в Дагестане мощность нижней и средней юры достигает 7 км), а в южном прогибе осадконакопление сопровождалось подводными вулканическими излияниями. С поздней юры оба прогиба развивались совершенно обособленно.. В северном формировались преимущественно карбонатные отложения, в южном - мощная толща флишевой формации.
Подобные условия осадконакопления сохранились и в меловом периоде. Центральная геоантиклиналь продолжала расширяться, прогибы все более обособлялись. В конце мелового периода произошло общее поднятие и регрессия моря.
По-иному шло развитие Малокавказской геосинклинальной системы. В юрском периоде на Малом Кавказе произошли резкие тектонические подвижки, возник ряд глубинных разломов, по которым из подкоровой части земной коры поднимался вверх магматический материал. По разломам отдельные крупные участки опускались,- другие поднимались. В образовавшемся почти широтном прогибе в морских условиях формировалась многокилометровая толща эффузивов основного состава.
В раннемеловую эпоху Малый Кавказ был охвачен поднятиями и представлял собой сушу, в позднем мелу произошли общие опускания и снова началось формирование мощных осадочно-вулканических формаций.
Мезозойская история других геосинклинальных систем Альпийско-Гималайской геосинклинальной области обнаруживает черты сходства с рассмотренной историей Кавказа. Все геосинклинальные системы этой области находились в мезозое на главном этапе геосинклинального развития - повсеместно происходили сложные движения земной коры, усложнялась тектоническая структура, накапливались мощные толщи осадочных и вулканических пород различного состава. В окраинных геосинклинальных прогибах, расположенных около платформенных массивов (Альпы, Большой Кавказ и др.), накапливались преимущественно песчано-глинистые и карбонатные осадки без вулканического материала. Во внутренних прогибах (Малый Кавказ и др.), наоборот, вулканогенные отложения преобладают.
Тихоокеанский геосинклинальный пояс
Тихоокеанский пояс окружает впадину Тихого океана в виде неравномерно расширяющейся и сужающейся полосы. В течение всей истории Земли этот пояс всегда был самой крупной геосинклинальной структурой. В мезозое в его пределах выделялось несколько геосинклинальных областей, которые примыкали непосредственно к впадине Тихого океана. Характерной особенностью развития этих областей было повсеместное проявление мезозойской складчатости, которая в разных геосинклинальных областях протекала не одинаково. В одних областях мезозойская складчатость привела к завершению геосинклинального режима в конце мезозоя (Кордильерская, Верхоянско-Чукотская); в других геосинклинальное развитие продолжалось и в кайнозойскую эру (Восточно-Азиатская, Андийская и др.).
Мезозойский орогенез вызвал значительные изменения в палеогеографической обстановке на территории Тихоокеанского пояса. Возникли обширные горные массивы, сократились площади морских бассейнов, сильно изменились очертания материков и морей.
Урало-Монгольский складчатый пояс
В начале мезозоя вся территория этого пояса представляла собой горную складчатую страну, в пределах которой геосинклинальный режим завершился в конце палеозойской эры. В течение мезозоя на всей площади пояса господствовали процессы размыва и лишь кое-где в изолированных впадинах накапливались континентальные отложения, которые в ряде участков являются угленосными. Морское осадконакопление наблюдалось в юрском и меловом периодах только в северо-западной части пояса. Оно было вызвано опусканиями в области современной Западно-Сибирской низменности и трансгрессией моря из Арктического бассейна. С мезозойскими морскими отложениями связаны здесь крупнейшие месторождения нефти и горючего газа.
История геологического развития древних платформ
В мезозойскую эру геологическая история Лавразии существенно отличалась от истории Гондваны. Платформенный массив Лавразия, в состав которого входили все древние платформы северного полушария, в триасе находился в континентальных условиях, а в юре и мелу крупные участки его территории испытали опускания и трансгрессию моря. Южный платформенный массив - Гондвана - в течение мезозоя испытал значительные преобразования. Он распался на части, между которыми возникли крупные океанические впадины.
Восточно-Европейская древняя платформа
Мезозойская история этой платформы протекала в достаточно спокойной обстановке. В триасовом периоде платформа была приподнятым участком земной коры и накопление континентальных аллювиальных и озерных отложений происходило в Московской, Польско-Литовской и Прикаспийской синеклизах. Главными источниками сноса обломочного материала были горные массивы Урала и Тимана. Только в южную часть Прикаспийской синеклизы проникало море с юга - из Средиземноморского пояса.
В начале юрского периода прогибание охватило небольшие участки на юге платформы и море проникло на территорию Донбасса. Морской бассейн значительно расширился в средней юре, он занимал обширную Украинскую синеклизу, образовавшуюся севернее Украинского щита и соединяющуюся на юго-востоке с Прикаспийской синеклизой, которая тоже была покрыта морем. В средней юре прогибания охватили западную и северо-восточную части платформы (Польско-Литовскую и Печорскую синеклизы). Повсеместно накапливались маломощные песчано-глинистые отложения. Продолжающиеся прогибания привели к расширению морского бассейна, и в поздней юре трансгрессия достигла максимума. В центральной части платформы соединились морские воды, трансгрессировавшие с юга, запада и северо-востока. Возник обширный морской бассейн, в котором все время существовали мелководные условия. На больших площадях этого бассейна накапливались однообразные темные глины с горизонтами песков и фосфоритов. Мощность верхнеюрских глин достигает всего нескольких десятков метров.
В начале мелового периода морской бассейн на платформе еще охватывал значительную территорию, но площадь его сокращалась. Происходило обмеление, и постепенно увеличивалась роль песчаного материала в отложениях. Связь с Средиземноморским и Арктическим бассейнами сохранялась почти до конца раннемеловой эпохи.
На рубеже раннего и позднего мела произошло осушение северной части Восточно-Европейской платформы и очертания морского бассейна на ней стали иными. Прогибания сохранились только в южной части платформы, где находился морской бассейн широтного направления, входивший в состав древнего океана, получившего название Тетис. Этот огромный океан существовал в центральной части восточного полушария еще в палеозойскую эру и в течение геологической истории менял свои очертания, то расширяясь, то сужаясь. В позднемеловую эпоху Тетис достиг максимальных размеров. Это было время крупнейшей в истории Земли морской трансгрессии, охватившей всю территорию Средиземноморского пояса и значительные площади Восточно-Европейской и Северно-Африканской древних платформ. На Восточно-Европейской платформе, да и в других участках Тетиса, резко изменился характер осадконакопления в позднемеловом бассейне по сравнению с более ранними эпохами. Резко возросла роль карбонатных осадков, повсеместно накапливались мергели, известняки, карбонатные глины и писчий мел.
В конце мезозойской эры территорию Восточно-Европейской платформы охватили поднятия и море ушло за ее пределы.
Сибирская древняя платформа
В течение мезозойской эры большая часть этой платформы представляла собой приподнятую область. Море заливало только северную и северо-восточную окраины.
В триасовом периоде в западной части платформы в Тунгусской синеклизе продолжалось образование мощной трапповой формации, которое началось еще в конце пермского периода. Трапповый вулканизм достиг максимума в начале триаса, вулканические извержения магмы основного состава происходили в континентальных условиях. На геологической карте Сибирской платформы хорошо видно распространение триасовых сибирских траппов в Тунгусской синеклизе на площади более 1,5 млн. км 2 . В юрский и меловой периоды на Сибирской платформе возникли и формировались две крупные синеклизы; на севере - Хатангская и на северо-востоке - Ленско-Вилюйская. Их контуры отчетливо выступают на геологической карте. В Хатангской синеклизе накапливались преимущественно морские, а в Ленско-Вилюйской - континентальные отложения. Большое значение имеют верхнеюрские и нижнемеловые отложения Ленско-Вилюйской синеклизы, представляющие собой мощную угленосную серию с колоссальными запасами каменного и бурого угля (2648 млрд. т). В юрский период угленакопление происходило и на юго-западе Сибирской платформы. Здесь в сравнительно небольшой Канской и Иркутской впадине накопилась угленосная формация большой мощности. Особенно велики запасы угля в Канском бассейне (1220 млрд. т), в Иркутском они значительно скромнее (87 млрд. т).
Литература
1. Аугуста И., Буриан З. По путям развития жизни. – Прага, 1971
2. Гангиус А. Через горы времени. – М., 1993
3. Муратов М.В. Происхождение материков и океанических впадин. – М., 1995
4. Немков Г.И. Левицкий Е.С. и др. Краткий курс палеонтологии. – М., 1988