Позвоночные объединены общностью морфофизиологической организации. Во всех системах органов этих животных можно проследить черты преемственных изменений в связи с эволюционным преобразованием гомологичных органов. Ниже излагается общий план строения, функционирования и закладки в онтогенезе отдельных систем органов.
Кожные покровы представляют собой весьма важную в функциональном отношении систему. Кожа непосредственно контактирует с внешней средой и испытывает прямое ее воздействие. Кожа и мускулы оформляют тело животного с поверхности, придают ему форму и удерживают все внутренние органы. Кожные покровы защищают тело от внешних механических и химических повреждений, воздействия температуры, иссушения, проникновения микробов. Кожа принимает участие в теплорегуляции, газообмене и выведении продуктов распада. Производные кожи могут принимать участие в формировании органов передвижения (копыта), служить для хватания (когти), нападения и защиты (рога, иглы и др.), полета (складки), плавания (перепонки).
Кожа содержит рецепторы органов осязания, в ней много желез разного назначения (слизистые, жировые, пахучие, потовые и пр.).
Для кожи позвоночных характерна двуслойность. Наружный слой - эпидермис - имеет эктодермальное происхождение. Он всегда многослоен. Нижний слой его пожизненно остается живым и деятельным и продуцирует новые слои клеток. Верхние слои эпидермиса составлены обычно уплощенными клетками, которые у наземных позвоночных ороговевают, отмирают и постоянно
слущиваются. Эпидермис дает начало роговым производным кожи - роговым чешуям, перьям, волосяному покрову, когтям, копытам, полым рогам. В эпидермисе развиваются разнообразные кожные железы.
Внутренний волокнистый слой кожи - кориум , иначе именуемый кутисом , или собственно кожей, развивается из мезодермального зачатка - кожного листка сомита. Слой кориума толст, он составляет основную часть кожи, обладающую большой прочностью. В кориуме развиваются разнообразные окостенения в виде чешуи рыб, покровных костей, образующих так называемый кожный скелет (в отличие от хондрального скелета). За счет собственно кожи развиваются также костные рога оленей. В нижней части кутиса накапливается подкожный слой жира.
Мускулатура . Располагающийся под кожей слой мышц представляет собой основную массу мускулатуры, именуемую мускулатурой тела , или соматической . Она обеспечивает животным возможность движения в среде и состоит из поперечно-полосатой мышечной ткани. У низших позвоночных, как и у бесчерепных, мускулатура имеет сегментированный характер. У высших позвоночных в связи с общим усложнением телодвижений, с развитием конечностей сегментация нарушается, и туловищная мускулатура группируется, оформляя такие части тела, как торс, голову, органы движения.
Кроме соматической мускулатуры у позвоночных есть мускулатура кишечника и некоторых других внутренних органов (сосудов, каналов). Эта мускулатура носит название висцеральной. Она сложена гладкой мышечной тканью и обеспечивает, в частности, движение пищи в кишечнике, сокращение стенок кровеносных сосудов.
Туловищная мускулатура эмбрионально возникает из внутреннего листка миотома (см. развитие ланцетника), т.е. спинного отдела мезодермы. Висцеральная мускулатура - это производное боковой пластинки, т.е. брюшного отдела мезодермы.
Внутренний скелет - это опорная основа тела позвоночного животного. Кроме того, скелет участвует в движении тела, осуществляет защиту внутренних органов.
Топографически скелет позвоночных может быть разделен на осевой, висцеральный, скелет поясов конечностей и свободных конечностей.
Осевой скелет в первоначальном виде представлен хордой , окруженной толстой соединительнотканной оболочкой. Последняя охватывает не только хорду, но и лежащую над ней нервную трубку. Хорда развивается из зачатка спинной стороны первичной кишки, т.е. имеет энтодермальное происхождение.
Рис. 9. Образование позвонка (поперечные разрезы):
1 - зачаток нижней дуги; 2 - зачаток верхней дуги; 3 - ребро, 4 - гемальный отросток, 5 - боковой отросток позвонка, 6 - верхняя дуга
У большинства позвоночных хорда вытесняется и замещается хрящевым или костным скелетом. Хрящевой и костный скелеты развиваются как производные указанной выше соединительнотканной (мезодермальной по происхождению) оболочки. Оболочка эта является, таким образом, скелетогенной.
В осевом скелете различают позвоночный столб и мозговой череп . При развитии позвонков первоначально закладываются метамерно расположенные парные хрящи, прилегающие к поверхности хорды. Это зачатки верхних и нижних дуг позвонков (рис. 9,1). Разрастание и смыкание наружных концов верхних дуг приводит к формированию спинномозгового канала , в котором располагается нервная трубка (рис. 9, II). Нижние дуги смыкаются в хвостовом отделе (у рыб) и ограничивают гемальный канал, где проходят спинная аорта и хвостовая вена. В результате смыкания внутренних концов верхних и нижних дуг образуются тела позвонков, внутри них и между ними в той или иной мере может сохраняться хорда (рис. 9, III). В туловищном отделе к отросткам нижних дуг позвонков причленяются ребра.
Мозговой череп , или черепная коробка, закладывается в виде двух пар хрящей, лежащих под зачатком головного мозга. Задняя их пара - парахордалии - располагается по бокам переднего конца хорды; передняя пара - трабекулы - впереди от нее. В них преобразуются зачатки первых позвонков и отчасти дуги висцерального скелета. Разрастание и смыкание парахордалии и трабекул приводят к образованию основной пластинки черепа, подстилающей головной мозг. Одновременно вокруг закладывающихся органов чувств (обоняния, зрения, слуха) возникают хрящевые капсулы. Они располагаются несколько выше уровня основной
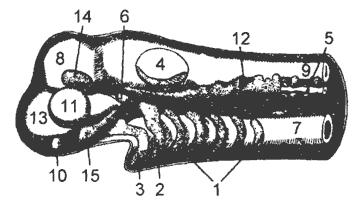
Рис. 10. Развитие хрящевого черепа акулы:
1 - жаберные дуги; 2 – подъязычная дуга; 3 - челюстная дуга; 4 – слуховая капсула; 5 - хорда; 6 - гипофиз; 7 -кишка; 8 - средний мозг; 9 – спинной мозг; 10 - ноздря; 11 - глаз; 12 - парахордалии; 13 - первичный передний мозг; 14-глазничные хрящи; 15 – трабекулы
пластинки черепа и закрывают головной мозг с боков. В последующем развитии капсулы органов чувств соединяются и срастаются с основной пластинкой черепа. При хрящевом состоянии черепа полной крыши у мозговой коробки не возникает. Остающиеся между перемычками крыши черепа отверстия - фонтанели - затянуты соединительнотканной перепонкой. Сплошная крыша черепа возникает лишь в связи с образованием накладных (кожных) костей (лобных, теменных).
Таким образом, мозговой череп возникает в связи с развитием головного мозга и органов чувств как защитное их образование.
Висцеральный скелет филогенетически формируется независимо от мозгового черепа (рис. 10). Его закладка происходит в соединительной ткани вблизи передней части пищеварительной трубки. Первоначально висцеральный скелет - это ряд многочисленных однообразных дуг, расположенных между жаберными щелями. Они служат опорой дыхательному аппарату.
С последующим преобразованием висцерального скелета связано приобретение позвоночными таких органов, как верхние и нижние первичные челюсти, среднее ухо, дно мозгового черепа, гортань.
Понятие "висцеральный скелет" рассматривается в основном применительно к низшим позвоночным животным. У высших оно заменяется понятиями "висцеральный череп", "лицевой череп".
Скелет поясов конечностей и свободных конечностей . У позвоночных животных различают конечности непарные и парные. В свою очередь парные конечности могут быть или плавниками, или конечностями наземного типа.
Скелет непарных конечностей - спинного, хвостового, анального плавников - состоит из ряда хрящевых или костных лучей, не связанных с другими частями скелета.
Скелет парных конечностей подразделяется на скелет поясов конечностей и скелет свободной конечности. Пояса конечностей всегда располагаются внутри тела животного. Скелет свободной конечности у позвоночных бывает двух типов: плавник рыб и

Рис. 11. Схема скелетов позвоночных:
I - скелет рыбы, II - скелет наземного позвоночного
пятипалая конечность наземных позвоночных. В первом случае скелет представлен несколькими рядами хрящиков или косточек, которые перемещаются относительно пояса как единый рычаг. Скелет пятипалой конечности состоит из ряда рычагов, способных перемещаться и совместно относительно пояса конечностей, и отдельно - один относительно другого (рис. 11). Закладка скелета конечностей происходит в соединительнотканном слое кожи.
Органы пищеварения . Система пищеварительных органов представлена трубкой, начинающейся ротовым отверстием и заканчивающейся анальным отверстием. Пищеварительный тракт формируется из энтодермальной трубки гаструлы (см. развитие ланцетника). В связи с этим эпителий пищеварительного тракта является энтодермальным. Только в области ротового и анального отверстий энтодермальный эпителий незаметно переходит в эктодермальный. Это связано с впячиванием стенок тела (а следовательно, и эктодермы) при образовании указанных выше отверстий.
Пищеварительный тракт подразделяется на следующие основные отделы: 1) ротовая полость , служащая для принятия пищи; 2) глотка - отдел, всегда связанный с органами дыхания: у рыб в глотку открываются жаберные щели, у наземных позвоночных в глотке располагается гортанная щель; глотку справедливо называют дыхательным отделом пищеварительной трубки; 3) пищевод , 4) желудок - расширение кишечного тракта, имеющее в некоторых случаях весьма сложное устройство; 5) кишечник , в типичном случае подразделяющийся на переднюю, или тонкую, среднюю, или толстую, и заднюю, или прямую, кишку. Морфологическое усложнение кишечного тракта в ряду позвоночных идет по пути
его удлинения и дифференцировки на отделы. В пищеварительную трубку открываются протоки трех видов пищеварительных желез: слюнных, печени, поджелудочной .
Слюнные железы - приобретение наземных позвоночных. В них преобразуются слизистые железы ротовой полости. Секрет их смачивает пищу и способствует расщеплению углеводов.
Печень и поджелудочная железа развиваются путем выпячивания переднего отдела эмбриональной кишки. Печень возникает из слепого выроста брюшной стенки кишечника (см. печеночный вырост кишки ланцетника). Протоки печени впадают в передний отдел тонкой кишки. Поджелудочная железа развивается из нескольких, чаще из трех, зачатков, представляющих собой также выросты кишечника. Эта железа, в отличие от печени, обычно не имеет вида компактного тела, и ее дольки рассеяны по брыжейке переднего отдела тонкой кишки.
Функции обеих указанных желез шире, чем только пищеварительные. Так, печень кроме выделения желчи, эмульгирующей жиры и активизирующей действие других пищеварительных ферментов, служит важным органом обмена веществ. Здесь нейтрализуются некоторые вредные продукты распада, накапливается гликоген. Ферменты поджелудочной железы расщепляют белки, жиры и углеводы. Одновременно поджелудочная железа служит органом внутренней секреции. Расстройство этой функции приводит к потере способности организма использовать сахар. В результате возникает тяжелое заболевание - диабет.
Органы дыхания позвоночных бывают двух типов - жабры и легкие, и у значительной части позвоночных существенное значение в дыхании имеет кожа.
Жаберный аппарат представляет собой систему парных, обычно симметрично расположенных, щелей, служащих для сообщения глотки с наружной средой. Передние и задние стенки жаберных щелей выстланы слизистой оболочкой, образующей пластинчатые выросты; выросты поделены на лепестки , носящие название жаберных . Каждая жаберная пластинка выше лепестков носит название полужабры . В промежутках между жаберными щелями (в жаберных перегородках) распалагаются висцеральные жаберные дуги (см. с. 27 о висцеральном скелете). Таким образом, каждая жаберная дуга связана с двумя полужабрами двух разных жаберных щелей.
Жаберные щели закладываются в виде системы парных энтодермальных выпячиваний, растущих из глотки кнаружи. Одновременно появляются эктодермальные впячивания наружных покровов. Зачатки растут навстречу друг другу и затем соединяются. Следовательно, жаберные щели имеют смешанное энто- и экто-дермальное происхождение. Жаберные лепестки Обычно
развиваются из эктодермального зачатка щели, и только у бесчелюстных они имеют энтодермальное происхождение.
Органы дыхания наземных позвоночных - легкие - в схеме представляют собой пару мешков, открывающихся в глотку через гортанную щель. Эмбрионально легкие возникают в виде выпячивания брюшной стенки глотки в задней части жаберного аппарата, т.е. имеют энтодермальное происхождение. На ранних стадиях развития эмбрионов зачатки легких напоминают пару внутренних (энтодермальных) жаберных щелей. Эти обстоятельства, а также общие для легких и жабр черты кровоснабжения и иннервации заставляют считать легкие гомологами задней пары жаберных мешков.
Кожа участвует в дыхании в случаях, когда в ней отсутствуют плотные роговые или костные чешуи, например у земноводных, голокожих рыб.
Функционально дыхательная система участвует в обогащении крови кислородом и в удалении углекислого газа. Через дыхательную систему у низших водных животных происходит сбрасывание аммиака. У теплокровных животных она участвует в процессах терморегуляции. Принцип работы дыхательной системы -обмен CO 2 и O 2 между потоками газа и крови, направленными противотоком навстречу друг другу.

Рис. 12. Схема кровообращения рыбы (I) и наземного позвоночного (II):
1 - жаберные артерии; 2 - сонная артерия; 3 - спинная аорта;4 - брюшная аорта; 5 - легочная артерия; 6 - кювьеров проток;7 - передняя кардинальная (яремная) вена; 8 - задняя кардинальная вена; 9 - задняя полая вена; 10 - печеночная вена; 11 - легочная вена;12 - воротная вена печени; 13 - подкишечная вена
Органы кровообращения . Кровеносная система у позвоночных, как и у бесчерепных, замкнутая. Закладывается кровеносная система из внутренних листков боковых пластинок (см. развитие ланцетника). Она состоит из сообщающихся между собой кровеносных сосудов, которые в грубой схеме могут быть сведены к двум стволам: спинному , где кровь течет от головы к хвосту, и брюшному , по которому она движется в обратном направлении. В отличие от бесчерепных (рис. 12) у позвоночных движение крови связано с деятельностью сердца.
Сердце представляет собой толстостенный мускульный мешок, разделенный на несколько отделов - камер. Основными отделами сердца являются предсердие, принимающее кровь, и желудочек, направляющий ее по телу. Число камер сердца различно у разных классов позвоночных.
Эмбрионально сердце возникает как расширение задней части брюшной аорты, которая в этом месте свертывается в изогнутую петлю. Передний отдел петли дает начало желудочку сердца, задний - предсердию .
Сердце имеет поперечно-полосатую мускулатуру, работающую в автоматическом режиме, и сокращение его не подчинено волевым импульсам. Размеры сердца связаны с интенсивностью его работы, и его размеры относительно размеров тела увеличиваются в ряду позвоночных (табл. 1).
Таблица 1
Сердечный индекс у позвоночных разных классов
Кровеносные сосуды подразделяются на две системы: артериальную , в которой кровь течет от сердца, и венозную , по которой кровь возвращается к сердцу. В процессе усложнения позвоночных наблюдается переход от животных, имеющих один круг кровообращения, к обладателям двух кругов кровообращения.
По своей природе кровь относится к соединительной ткани, проникающей в кровеносное русло из межклеточного пространства.
Кровь позвоночных состоит из бесцветной жидкости - плазмы, в которой находятся форменные элементы крови: красные кровяные тельца, или эритроциты, содержащие красящее вещество - гемоглобин, и белые кровяные тельца - лейкоциты. С эритроцитами связано окисление крови, так как они переносят кислород. Лейкоциты участвуют в уничтожении попавших в тело микроорганизмов. Кроме того, в крови есть тромбоциты, играющие важную роль в свертывании крови, а также прочие клетки. Масса крови в эволюционном ряду позвоночных увеличивается (табл. 2).
Таблица 2
Относительная масса крови к массе тела у позвоночных разных классов
(по Проссеру и Брауну, 1967; по Проссеру, 1978)
Кровеносная система полифункциональна. Она участвует в получении органами, тканями, клетками кислорода, органических и минеральных веществ, жидкостей и выносе продуктов распада, шлаков, углекислого газа, в переносе гормонов желез внутренней секреции и т.д.
Наряду с кровеносной системой у позвоночных есть другая, связанная с ней, сосудистая система - лимфатическая . Она состоит из лимфатических сосудов и лимфатических желез . Лимфатическая система незамкнута. Только крупные ее сосуды имеют самостоятельные стенки, в то время как их разветвления открываются в межклеточные пространства различных органов. Лимфатические сосуды содержат бесцветную жидкость - лимфу , в которой плавают лимфоциты, образующиеся в лимфатических железах. Движение лимфы обусловливается сокращением стенок, некоторых участков крупных сосудов (так называемых лимфатических сердец) и периодически изменяющимся давлением на сосуды различных органов.
Лимфатическая система служит посредницей в обмене веществ между кровью и тканями.
Нервная система . Функции нервной системы - восприятие внешних раздражений и передача возникающих возбуждений к клеткам, органам, тканям, а также объединение и согласование деятельности отдельных систем органов и организма в целом в единую функционирующую живую систему. Эмбрионально нервная система позвоночных возникает, так же как и у бесчерепных, в виде закладывающейся в эктодерме на спинной стороне зародыша полой трубки (рис. 13). В последующем происходит ее дифференцировка, приводящая к образованию: а) центральной нервной системы , представленной головным и спинным мозгом, б) периферической нервной системы , состоящей из нервов, отходящих от головного и спинного мозга, и в) симпатической нервной системы , состоящей в основе из нервных узлов, расположенных около позвоночного столба и связанных продольными тяжами.
Головной мозг представлен у позвоночных животных пятью отделами: передним, промежуточным, средним, мозжечком и продолговатым мозгом. Он закладывается эмбрионально в виде вздутия переднего отдела нервной трубки, которое вскоре делится на три первичных мозговых пузыря (рис. 14). В дальнейшем первый мозговой пузырь дает начало спереди переднему мозгу; задняя его часть преобразуется в промежуточный мозг. Из второго мозгового пузыря формируется средний мозг. Путем выпячивания
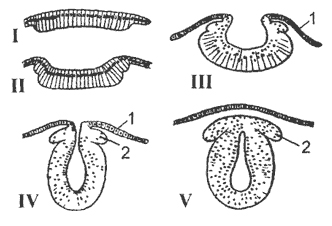
Рис. 13. Последовательные стадии развития центральной нервной системы позвоночных (схематические поперечные разрезы):
I-II дифференцировка эктодермы; III впячивание нервной пластинки, IV-V - обособление нервной трубки, 1 эпидермис; 2 ганглионарная пластинка
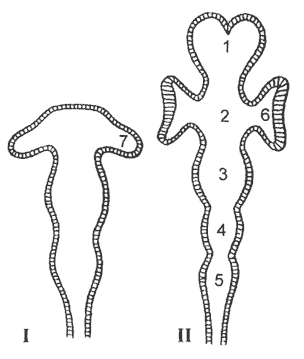
Рис. 14. Развитие отделов головного мозга (схема):
I - стадия трех пузырей (с глазными пузырями), II - стадия пяти отделов (с глазными бокалами), 1 - передний мозг, 2 - промежуточный мозг; 3 -средний мозг; 4 - мозжечок; 5 - продолговатый мозг; 6 - глазной бокал; 7 - глазной пузырь
крыши третьего мозгового пузыря формируется мозжечок, под которым располагается продолговатый мозг. Передний мозг, кроме того, подразделяется на левую и правую половины.
Одновременно с разрастанием и дифференцировкой головного отдела нервной трубки происходит соответственное преобразование невроцеля. Два его расширения в полушариях переднего мозга известны под названием боковых желудочков мозга. Расширенная часть невроцеля в промежуточном отделе мозга обозначается как третий желудочек , полость среднего мозга - как силъвиев водопровод , полость продолговатого мозга - как четвертый желудочек , или ромбовидная ямка (рис. 14). От головного мозга отходят 10 или 12 пар черепномозговых нервов .
Передний мозг имеет впереди два симметрично расположенных выступа, от которых отходит первая пара головных нервов - обонятельные (I). От дна промежуточного мозга отходят зрительные нервы (вторая пара головных нервов, II).
На крыше промежуточного мозга развиваются два сидящих на ножках выступа: передний - теменной орган и задний - эпифиз .
От дна промежуточного мозга отрастает непарный выступ - воронка , к которой прилегает сложное по строению и функции образование - гипофиз . Передний отдел гипофиза развивается из эпителия ротовой полости, задний - из мозгового вещества. Там же расположен гипоталамус .
Крыша среднего мозга образует парные вздутия - зрительные доли (бугры). От среднего мозга отходит третья пара головных нервов (глазодвигательные , III). Четвертая пара головных нервов (блоковые , IV) отходит на границе между средним и продолговатым мозгом, все остальные головные нервы (V - X - XII) отходят от продолговатого мозга.
Спинной мозг не отграничен резко от продолговатого мозга. В центре спинного мозга (по главной оси органа) сохраняется невроцель, известный у позвоночных под названием спинномозгового канала .
От спинного мозга метамерно (по числу сегментов) отходят спинномозговые нервы. Они начинаются двумя корешками: спинным - чувствующим и брюшным - двигательным. Эти корешки вскоре по выходе из спинного мозга сливаются, образуя спинномозговые нервы, которые затем вновь делятся на спинную и брюшную ветви.
Органы чувств . Эта группа органов возникает как производные разных частей зародыша и на разных этапах его развития. Это органы обоняния, зрения, слуха, вестибулярный аппарат, органы боковой линии, органы вкуса, осязания, специфические органы, воспринимающие магнитное поле Земли, электрические поля, тепловые излучения и пр.
Органы обоняния . Предполагают, что обоняние - одна из самых древних функций мозга. Органы обоняния закладываются как утолщение эктодермы одновременно с нервной пластинкой. Параллельно формируется скелет обонятельных капсул, которые входят в состав мозгового черепа. Поначалу обонятельные капсулы сообщаются только с внешней средой и имеют наружные ноздри. Впоследствии в связи с наземным существованием ноздри становятся сквозными.
Органы зрения также принадлежат к древним чувствующим органам. Светочувствительная рецепция возникает на очень раннем этапе эволюции хордовых (вспомним ланцетника) и формируется в раннем эмбриогенезе. Органы зрения позвоночных подразделяют на парные и непарные. И те и другие есть выросты промежуточного мозга. Парные глаза закладываются как выросты боковых частей промежуточного мозга, непарные - как последовательно расположенные в крыше промежуточного мозга (эпифиз и теменной орган). Закладка парных глаз сопровождается формированием около них зрительных капсул, входящих в состав мозгового черепа (рис. 15, 16).
Органы слуха имеют у позвоночных животных сложное происхождение. Наиболее рано в эволюции формируется внутреннее ухо, которое

Рис. 15. Последовательные стадии развития глаза:
1 - промежуточный мозг; 2 - эпидермис, формирующий хрусталик; 3 - начало формирования глазного бокала; 4 - ножка глазного бокала; 5 - хрусталик; 6 -роговица; 7 - сетчатка; 8 - пигментная оболочка; 9 - клетки мезодермы, формирующие сосудистую оболочку и склеру
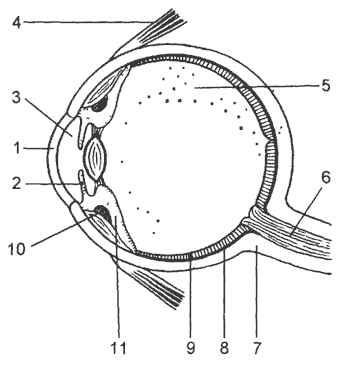
Рис. 16. Сагиттальный разрез глаза человека:
1 - роговица; 2 - радужина; 3 -передняя камера; 4 - одна из мышц, перемещающих глаз; 5 - стекловидное тело; 6 - зрительный нерв; 7 - склера; 8 - сосудистая оболочка; 9 - сетчатка; 10 - ресничная мышца; 11 - ресничное тело
закладывается в эктодерме зародыша, углубляется в виде ямки и оформляется как слуховой пузырек , лежащий в слуховой капсуле. Слуховой пузырек делится перетяжкой на две части. Верхний отдел превращается в вестибулярный аппарат. Это орган равновесия. Он позволяет ощущать положение тела в трехмерном пространстве Земли. Этот орган - 3 полукружных канала во внутреннем ухе (рис. 17, 18). Нижний отдел слухового пузырька представляет собой собственно внутреннее ухо - слуховой мешочек .
Среднее и наружное ухо формируются на поздних этапах возникновения позвоночных животных в связи с выходом на сушу.
Органы боковой линии свойственны только первичноводным позвоночным, закладываются также в эктодерме. Они представляют собой желобки, тянущиеся

Рис. 17. Последовательные стадии развития внутреннего уха:
I - слуховая плакода, II - ямка, III и IV - пузырек в разрезе; V и VI - образование полукружных каналов; 1 - зачаток улитки; 2 - полукружный канал; 3 -эндолимфатический проток; 4 - круглый мешочек; 5 - овальный мешочек
по бокам головы и вдоль тела. Эти желобки могут быть прикрыты (или нет - у голокожих) костной чешуей. Органы боковой линии воспринимают легкие движения и колебания воды вблизи от источников колебаний: скорость и направление течений, движений собственного тела и наличие предметов на пути движения животного в воде. Они представляют собой сейсмосенсорную систему.
Органы вкуса закладываются в энтодерме и воспринимают вкус потребляемой пищи в диапазоне: сладкая, горькая, кислая, соленая. Они располагаются на вкусовых сосочках в пределах ротовой полости.
Органы осязания . Чувствующих клеток не имеют, а представляют собой разветвленные в коже нервные окончания, которые и воспринимают предметы среды на ощупь.
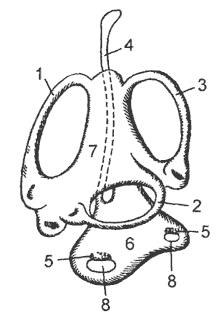
Рис. 18. Схема перепончатого слухового лабиринта:
1, 2, 3 - передний, наружный и задний полукружные каналы, 4 - эндолимфатический проток, 5 - слуховые пятна, 6 - круглый мешочек, 7 овальный мешочек, 8 – отолиты

Рис. 19. Боуменова капсула с сосудистым клубком:
1 - приносящий кровеносный сосуд, 2 - выносящий кровеносный сосуд,3 - боуменова капсула, 4 - сосудистый клубок, 5 - первичная моча,6 - почечный каналец, 7 - воронка почечного канальца (нефростом),открывающаяся в полости тела

Рис. 20. Схема мочеполовой системы позвоночных:
I (самец) и II (самка) - акуловые и амфибии, III (самец) и IV (самка) – рептилии и птицы, V (самец) и VI (самка) - млекопитающие, 1 - пронефрос (предпочка);2 - семенник; 3 - семявыносящий проток, 4 - мезонефрос (первичная почка),5 - задняя кишка, 6 - мочевой пузырь, 7 - клоака, 8 - воронка, 9 - яичник,10 - мюллеров канал, 11 - придаток семенника (остаток передней части мезо-нефроса), 12 - семяпровод, 13 - метанефрос (вторичная почка), 14-вторичный мочеточник, 15-рудимент мезонефроса, 16-яйцевод, 17-яйцеклетка, 18-белок, выделяемый железами стенки яйцевода, 19 - матка, 20 -зародыш в матке; 21 - влагалище, 22 - половой синус, 23 - предстательные железы; 24 – семенной пузырек, 25 - семяпровод, 26 - придаток семенника, 27 - промежность;28 - рудимент мюллерова канала, 29 - канал мезонефроса (первичной почки); 30 - копулятивный орган (penis), 31 - анальное отверстие
Органы выделения . У всех позвоночных выделительные органы представлены почками , предназначенными для выведения из тела излишков воды, минеральных солей и продуктов распада азотистого обмена в виде мочевины или мочевой кислоты. Они имеют мезодермальное происхождение, закладываясь на внешней стенке сомитов. Однако строение и механизм функционирования почек у разных групп позвоночных не одинаковы. В процессе эволюции позвоночных животных происходит смена трех типов почек: головная , или предпочка (пронефрос), туловищная , или первичная, почка (мезанефрос ) и тазовая , или вторичная, почка (метанефрос ). Разные типы почек имеют разные принципы процесса выделения: выделение из полости тела, смешанное выделение (из полости тела и из крови) и, наконец, только из крови. Одновременно происходят изменения в механизме реабсорбции воды. Выделение воды и растворенных в ней продуктов белкового обмена из полости тела оказывается возможным благодаря множеству воронковидных нефростом, открывающихся в полость тела. Выделение из крови происходит через мальпигиевы тельца почек. Выводные каналы из почек называются волъфовы каналы , их сменяют мочеточники . У большинства позвоночных есть мочевой пузырь . У первичноводных возможно сбрасывание аммиака через жабры.
Половые органы . Половые железы позвоночных - яичники у самок и семенники у самцов - как правило, парные. Они развиваются из отдела мезодермы в месте подразделения этого зачатка на сомит и боковую пластинку.
Первоначально (у бесчелюстных) половые железы не имели выводных протоков и половые продукты выпадали через разрывы стенок гонад в полость тела, откуда выводились в наружную среду через специальные поры. Впоследствии возникли половые пути, которые у самцов связаны с выделительными органами (вольфов канал). А у самок в качестве яйцевода функционирует мюллеров канал , который сохраняет связь целома с внешней средой.
Эволюция систем органов
«Ничто в биологии не
имеет смысла иначе
как в свете эволюции»
Ф.Г. Добржанский
Режим эволюции генов
Хрящевые рыбы
У хрящевых рыб (акулы, скаты) в сердце развивается четвертая дополнительная камера – артериальный конус. Внутри конуса имеется несколько пар клапанов, препятствующих возвращению крови в желудочек. Кроме этого, эта новообразованная мышечная ткань сердца сглаживает резкое пульсирующее движение крови и делает его более равномерным. Из артериального конуса кровь направляется в брюшную аорту и далее в жабры. У большинства хрящевых рыб жабры расположены на четырех хрящевых жаберных дугах и на подъязычной дуге, и соответственно имеется пять пар приносящих жаберных артерий. У животных с легочным типом дыхания некоторые жаберные артерии редуцировались (редукция – полное исчезновение органа или ткани), а часть преобразовалась в крупные кровеносные сосуды. Магистральные вены впадают в пару кювьеровых протоков, а затем в венозный синус, как у ланцетника. Появляется еще одна капиллярная сеть – воротная система почек. Количество эритроцитов у хрящевых рыб значительно больше, чем у круглоротых, что способствует снабжению тканей кислородом. Впервые появляется селезенка – орган иммунитета и место образования форменных элементов крови.
У костных (большинства пресноводных) рыб артериальный конус подвергся редукции, но расширилось основание аорты (луковица аорты). Количество эритроцитов значительно больше, чем у хрящевых рыб. Имеются сведения, что сердце костистых рыб не имеет симпатической иннервации, а снабжено только ветвями блуждающего нерва, который регулирует сердечные сокращения.
Двоякодышащие рыбы
Среди современных костных рыб представляют интерес двоякодышащие рыбы; они являются родственниками кистеперых рыб, от которых произошли наземные позвоночные. Морфофизиологической особенностью рыб, дышащих жабрами и легкими, является решение проблемы рационального использования обоих органов газообмена в воде и воздухе. Легкие у них сформировались позади глотки, т.е. позади жабер. В легкие кровь поступает по легочной артерии, начинающейся от задней верхней жаберной артерии. Оксигенированная (обогащенная кислородом) кровь возвращается из легких в сердце.
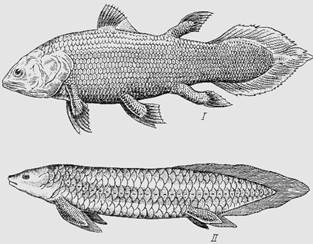
Рис. Кистеперые и двоякодышащие рыбы:
I – латимерия; II – неоцератод
У двоякодышащих рыб сердце состоит из венозного синуса, получающего венозную кровь, предсердия, желудочка и артериального конуса. В предсердии, желудочке и артериальном синусе имеются неполные внутренние перегородки. Венозный синус, несущий дезоксигенированную кровь от всего тела, впадает в правую половину предсердия, а в левую часть предсердия впадают легочные вены, несущие богатую кислородом кровь из легких. Из левой половины предсердия кровь попадает в левую половину желудочка и через артериальный конус – в нижнюю часть укороченной брюшной аорты и далее к двум передним парам приносящих жаберных артерий: затем кровь поступает по сонным артериям в голову. Из правой половины желудочка дезоксигенированная кровь направляется в верхнюю часть брюшной аорты и по ней же – к двум задним парам жаберных артерий.
Таким образом, у двоякодышащих рыб появляется второй легочный (малый) круг кровообращения. Чтобы не допустить проникновения воды в легкие, дыхательные пути перекрываются за счет сокращения мышц гортани; Вовремя закрыть эту щель позволяют нервные импульсы, посылаемые центральным генератором ритма в стволе мозга. Такая система регуляции имеется и у головастиков лягушки, которые дышат жабрами. Отголоски этой реакции сохранились в виде икоты, которая нередко возникает у человека. Икота представляет собой особую форму дыхательных движений, когда за резким вдохом следует быстрое перекрывание голосовой щели. Резкое перекрывание дыхания вызывает звук похожий на «ик». Приступы икоты могут продолжаться годами. (У Чарлза Осборна наблюдался с 1922 по 1991 год, и он занесен в книгу рекордов Гиннесса).
Следует подчеркнуть, что икота – это генетический физиологический признак. Причем очень древний. По меньшей мере, ему 370 миллионов лет. В то время не только человека, но и рептилий, птиц и млекопитающих не было на Земле. Таким образом, икота восходит своими корнями в далекое прошлое и свидетельствует о нашем эволюционном родстве с рыбами и амфибиями. Это называется эволюционной наследственностью. Икают не только дети и взрослые люди, но и эмбрионы, достигшие возраста двух месяцев. Назначение этого древнейшего рефлекса у эмбрионов млекопитающих двояко: 1) не допустить попадание околоплодных вод в развивающиеся легкие; 2) подготовиться к сосанию молока, при котором также необходимо перекрывать вход в гортань.
У наземных позвоночных произошли значительные изменения в строении кровеносной системы, наметившиеся у двоякодышащих рыб. У всех четвероногих брюшная аорта (частично замещенная сердцем), разделена на несколько обособленных артериальных стволов, начинающихся от сердца. Разделение брюшной аорты произошло по-разному у предков современных земноводных, пресмыкающихся и млекопитающих.
Земноводные (Амфибии)
Кровеносная система земноводных прогрессивно изменилась по сравнению с той же системой рыб, в связи выходом на сушу и заменой жаберного дыхания на легочное. У современных земноводных от артериального конуса сердца начинаются три пары сосудов: сонные артерии, дуги аорты, (которые на спинной стороне сливаются, образуя спинную аорту) и легочные или кожно-легочные артерии. По сонным артериям амфибий кровь течет к голове; по дугам аорты – в спинную аорту, а от неё к различным органам тела амфибий; легочные (или кожно-легочные) артерии обеспечивают кровоснабжение органов дыхания.
Сердце земноводных состоит из 3 основных отделов (левого и правого предсердий, желудочка) и двух дополнительных (венозного синуса, и артериального конуса). У земноводных, например, у лягушки, в полость желудочка от его стенок выступают мышечные гребни, которые разделяют его на отсеки. Наличие гребней частично препятствует смешению оксигенированной и дезоксигенированной крови. При легочном дыхании в артериальный конус сначала поступает кровь с низким содержанием кислорода и направляется в ближайшие к сердцу кожно-легочные артерии; вторая порция смешанной крови направляется в дуги аорты, и третья (наиболее обогащенная кислородом) кровь поступает по сонным артериям к голове. Если животное находится под водой и дышит кожей, то крови больше направляется к коже.
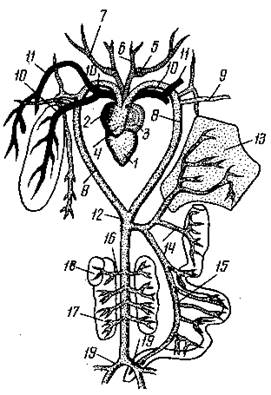
Рис. Схема артериальной системы лягушки: 1 – желудочек сердца; 2 – правое предсердие; 3 – левое предсердие; 4– артериальный конус; 5 -7 – ветви сонных артерий; 8 – дуги артерий; 9 – подключичная артерия; 10 – легочная артерия; 11 – большая кожная артерия; 12 – спинная аорта; 13 – печень; 14 – желудочная артерия; 15 – кишечная артерия; 16 – почечная артерия; 17 – почка; 18 – семенник; 19 – подвздошная артерия.
(Следует отметить, что у рыб все дуги жаберных артерий связаны друг с другом над глоткой корнями спинной аортой; такая связь сохраняется у некоторых четвероногих животных в виде сонных протоков между сонными артериями и дугами аорты, а также боталловых протоков между дугами аорты и легочными артериями.) У человека только один боталлов проток соединяет левую легочную артерию с дугой аорты во внутриутробном развитии. Незаращение боталлова протока у новорожденного является атавистическим врожденным пороком сердца.
В школьных учебниках для «убедительной» иллюстрации развития системы от простой к более сложной организации, даются упрощенные представления о строении сердца у рыб, амфибий и других позвоночных. Наблюдения показывают, что строение и регуляция сердечнососудистой системы амфибий являются довольно сложными, поскольку эти животные обитают в двух средах – водной и воздушной. К этому следует добавить, что у некоторых рыб в сердечной мышце имеются стволовые клетки, которые участвуют в регенерации миокарда. Этих клеток нет в миокарде у человека, поэтому на месте инфаркта образуется рубцовая ткань, препятствующая работе сердца.
Пресмыкающиеся
У пресмыкающихся имеет место ключевой ароморфоз – развитие мощного легочного насоса – грудной клетки. В связи с этим необходимость в кожном дыхании отпадает и появляется возможность разделения артериального и венозного потока крови.
Терморегуляция у рептилий несовершенна, поэтому для них характерна так называемая гелиотермия – обогревание тела в лучах солнца. При «приеме солнечных ванн» необходимо направлять больше крови к кожным покровам и меньше в легкие, а при активной деятельности – в легкие. В соответствии с этими потребностями, регулируется движение крови в большом и малом кругах кровообращения.
Брюшная аорта у предков рептилий разделилась на три артериальных ствола, начинающихся от сердца: левую и правую дуги аорты, сливающиеся с в спинную аорту и общий ствол легочных артерий. Правая дуга аорты получает из желудочка сердца наиболее чистую артериальную кровь; в связи с этим сонные и подключичные артерии, несущие кровь к голове и передним конечностям, у рептилий начинаются от правой дуги аорты. Строение венозного отдела кровеносной системы не претерпело существенных изменений по сравнению с амфибиями.
Сердце у большинства пресмыкающихся (ящерицы, змеи, черепахи), трехкамерное и состоит из 2-х полностью разделенных предсердий и одного желудочка. В левое предсердие впадают легочные вены, несущие оксигенированную кровь. В правое предсердие – три полые вены. В желудочке пресмыкающихся (кроме крокодилов) имеется неполная перегородка. Перегородка располагается не в вертикальной (как межпредсердная), а в горизонтальной плоскости, и поэтому разделяет желудочек на спинной и брюшной отделы. Оба предсердия открываются в спинной отдел желудочка. Следует отметить, что предсердия у рептилий (в отличие от птиц и млекопитающих), сокращаются не одновременно (поочередно). Первым сокращается правое предсердие, и венозная кровь попадает в брюшной отдел желудочка, а далее направляется в легкие для обогащения кислородом. Затем сокращается левое предсердие, и кровь заполняет спинной отдел желудочка. От этого отдела начинаются правая и левая дуги аорты, которые перекрещиваются. Во время систолы желудочка перегородка полностью отделяет брюшной отдел от спинного, так что в обе дуги аорты поступает артериальная кровь, а в легочные артерии – венозная.
В утренние часы у рептилий температура тела снижается, и животные вынуждены переходить в режим инсоляции – обогреву на солнце. (Следует заметить, что температурная кривая у человека подчиняется такому же ритму как у рептилий: утром снижается, а к вечеру – повышается). Во время инсоляции просвет общего ствола легочных артерий сужается, и часть крови из брюшного отдела желудочка направляется в спинной отдел, где происходит её смешение. Начало левой дуги аорты расположено правее и ниже начала правой, поэтому смешанная кровь поступает в левую дугу аорты, а в правую – оксигенированная кровь. От правой дуги аорты начинается сонные и подключичные артерии, поэтому голова и передние конечности рептилий получают только артериальную кровь.
Необходимо обратить внимание на тот факт, что горизонтальная перегородка в сердце рептилий является не удачным (инадаптивным) новообразованием, так как при горизонтальном полном структурном разделении желудочка кровь не смогла бы попадать в легочный артериальный ствол.
Настоящее четырехкамерное сердце развилось независимо в трех эволюционных линиях: у крокодилов, птиц и млекопитающих. Это считается одним из ярких примеров параллельной эволюции.
Параллельный эволюцией называют независимое появление похожих признаков у организмов разных таксонов (видов, родов, семейств, классов и др.). В параллелизме отчетливо проявляется закономерный, т.е. неслучайный характер эволюции. Это явление свидетельствует, что переход на более высокий уровень организации происходит не в одной крупной группе организмов, а в нескольких, развивающихся параллельно. При этом отдельные признаки, из которых складывается ароморфоз (усложнение организации), могут проявляться одновременно, а иногда и в разное время. Таким образом, прогрессивные признаки могут постепенно накапливаться до тех пор, пока в одной (или немногих) группе они не сформируются все вместе.
У крокодилов, относящихся к высшим рептилиям, произошла существенная перестройка сердца – образовалась вертикальная (сагиттальная) перегородка, полностью отделившая правый желудочек от левого. Причем от левого желудочка начинается правая (огибающая сердце справа) дуга аорты, а от правого – левая (огибающая сердце слева) дуга аорты и общий ствол легочных артерий. В том месте, где правая и левая дуги аорты перекрещиваются, имеется паниццево отверстие, через которое возможно смешение артериальной и венозной крови. Левая дуга аорты у крокодилов (а, так же, птиц, являющихся их родственниками), не сливается с правой дугой в спинную аорту, как у остальных рептилий, а продолжается в чревную артерию, снабжающую кровью кишечник. Спинную аорту образует лишь правая дуга. При нахождении крокодила под водой поток крови к легким резко ограничивается и венозная кровь из левой дуги аорты сбрасывается через паниццево отверстие в правую дугу аорты, где происходит ее смешение.



Строение сердца крокодила (RV – правый желудочек, LV – левый желудочек, FP – отверстие Паниццы, RA – правая аорта, LA – левая аорта, PA – лёгочная аорта)
// pharyngula.org/Газета.Ru
Рис. Артериальная система крокодила.
У рептилий клеточный материал венозного синуса, (функционирующий у амфибий), включается в стенку предсердий. Эта масса узловой ткани у птиц и млекопитающих образует синусный узел. Он находится в месте впадения верхней полой вены в правое предсердие. Ткань узла, имеющаяся только в сердце, стимулирует и регулирует сердечные сокращения. Она обладает некоторыми свойствами как мышечной, так, и нервной ткани. Через регулярные промежутки времени от синусного узла по мышце предсердия распространяется волна сокращения. Когда она доходит до атрио-вентрикулярного (предсердно-желудочкового) узла, импульс через пучок узловой ткани передается желудочкам.
Птицы
Птицы обособились от общего с крокодилами предка, поэтому кровеносная система птиц обнаруживает определенное сходство с таковой крокодилов. Появление четырехкамерного сердца у птиц и млекопитающих было важнейшим эволюционным событием, благодаря которому эти животные смогли стать теплокровными. Следует отметить, что у птиц появились более совершенные, чем у пресмыкающихся легкие и двойное дыхание, которое в достаточной степени обеспечивают организм кислородом. Такое прогрессивное преобразование дыхательной системы птиц обеспечило им теплокровность. Для птиц отпала необходимость в инсоляции и перераспределении крови в сосудах на суше и под водой. Левая дуга аорты у птиц полностью редуцировалась. Сердце птиц четырехкамерное, от сердца отходят два артериальных ствола: правая дуга аорты начинается от левого желудочка, а общий ствол легочных артерий – от правого желудочка. Таким образом, морфофизиологический прогресс (по А.Н.Северцову) достигнут не усложнением, а упрощением и рационализацией дыхательной и кровеносных систем. Воротная система почек у птиц отсутствует, кровоснабжение почек осуществляется почечными артериями, отходящими от спинной аорты.

Млекопитающие
У предков млекопитающих брюшная аорта разделилась не на три, как у пресмыкающихся, а на два артериальных ствола: общий ствол дуг аорты и общий ствол легочных артерий. Сердце млекопитающих четырехкамерное, как у птиц. Желудочек сердца разделился вертикальной перегородкой на левый (от которого начинается общий ствол дуг аорты) и правый (откуда берет начало легочный ствол). Впереди общий ствол дуг аорты разделяется на левую дугу аорты (продолжающуюся в спинную аорту) и безымянную артерию, переходящую в правую подключичную артерию. Сонные артерии начинаются симметрично: левая – от левой дуги аорты, правая – от безымянной артерии.
В эмбриональном развитии млекопитающих и человека рекапитулируют (повторяются) закладки сердца и основных кровеносных сосудов предковых классов. Сердце закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным. Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
Детальное изучение развития сердца у эмбрионов ящерицы и черепахи и сравнение его с имеющимися данными по амфибиям, птицам и млекопитающим показало, что ключевую роль в превращении трехкамерного сердца в четырехкамерное сыграли изменения в работе регуляторного гена Tbx5, который функционирует в изначально едином зачатке желудочка. Если Tbx5 эспрессируется (работает) равномерно по всему зачатку, сердце получается трехкамерным, если только с левой стороны - четырехкамерным.
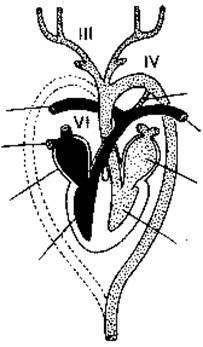

Расположение боталлова протока (схема): 1 - легочная артерия; 2 - правая ветвь легочной артерии; 3 - дуга аорты; 4 - боталлов (артериальный) проток.
Эволюция систем органов
«Ничто в биологии не
имеет смысла иначе
как в свете эволюции»
Ф.Г. Добржанский
Биологическая эволюция – необратимый процесс исторического развития живой природы. Определяется изменчивостью, наследственностью и естественным отбором. Сопровождается приспособлением организмов к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Эволюция (от лат. evolution – развертывание) в широком смысле – то же, что развитие; в более узком – представление о медленных, постепенных изменениях. Эволюция является ключевым моментом в устройстве и механизме мироздания. В связи с этим различают эволюцию неживой природы, биологическую и социально-культурную формы эволюции. Эволюционное мировоззрение позволяет увидеть общее в частностях и единство в многообразии.
В биологии под развитием понимают процесс тесно взаимосвязанных количественных (рост) и качественных (дифференцировка) преобразований особей с момента зарождения и до конца их жизни (онтогенез), а также и в течение всего времени существования жизни их видов на Земле (историческое развитие или филогенез).
Эволюция – твердо установленный научный факт. Она придаёт смысл, логику и стройность всему гигантскому массиву накопленных биологией знаний. Отвлечься от эволюции биолог может только на краткое мгновение, изучая, например, строение двадцать первой пары ножек у сороконожек, межклеточные контакты, строение нефрона почки человека, нейрона ЦНС, или другие структуры. Однако, выяснить как возникли, сложные структурно-функциональные единицы органов и сами органы, а также новые виды организмов помогает эволюционный (исторический) подход. Без эволюционного мировоззрения биология превращается в набор бессмысленных, разрозненных, необъясненных научных данных. Знания эволюции животных необходимы для понимания причин возникновения многих болезней и пороков развития. Сведения, полученные при исследовании организмов, находящихся на различных уровнях развития (вирусов, бактерий, протистов, червей, насекомых, рыб и других менее сложноорганизованных организмов) позволяют нам глубже понять, как работают человеческие органы, какие причины вызывают у нас болезни, и какие новые методы мы можем использовать для улучшения здоровья и продления нашей жизни.
Режим эволюции генов
Наиболее важным в эволюционном учении является вопрос о том, каким образом организмы приспосабливаются к внешней среде. Правильно ответить на этот кардинальный вопрос помогает биоинформатика. Ученые, работающие в этой передовой области (отрасли) генетики анализируют последовательности нуклеотидов в генах и порядок расположения аминокислот в белках. Основываясь на данных биоинформатики, японский биолог Мотто Кимура предложил теорию молекулярной эволюции. Согласно его взглядам, (получивших широкое признание), существует три режима накопления одиночных замен в генах.
Первый – нейтральная эволюция, когда мутации не изменяют структуру или функцию гена и соответствующего белка, Они характерны для участков ДНК, которые не слишком важны для выживаемости организма и не влияют на его приспособленность. (Такими участками, например, являются неинформативные участки гена – интроны, межгенные последовательности – спейсеры). В ходе эволюции такие мутации накапливаются в геномах с приблизительно постоянной скоростью, поэтому в молекулярной эволюции они служат эталоном времени. Процесс накопления таких замен даже назвали молекулярными часами по аналогии с атомными часами – эталоном времени в физике. (Используя, например, «молекулярные часы», ученые определили время расхождения ветвей от общего предка, приведших к шимпанзе и человеку).
Второй режим – стабилизирующий отбор, под действием которого обычно находятся особо важные гены или их участки; почти любая замена в них повредит продукт гена и уменьшит приспособленность организма. Скорость накопления мутаций при таком отборе тем ниже, чем важнее ген и чем меньше его функция связана непосредственно с внешней средой. Под действием стабилизирующего отбора часто находятся гены «домашнего хозяйства», кодирующие белки-ферменты энергетического обмена, белки-гистоны и мн. др.
Третий режим эволюции – движущий отбор. Он характеризуется увеличенной скоростью накопления мутаций, так как в новых условиях они повышают приспособленность организма, придавая генам какие-либо новые свойства (например, возрастает устойчивость микроорганизмов к антибиотикам, насекомых – к инсектицидам.
В эволюционном учении выделяют 4 основных направления эволюционной мысли: 1) ламаркизм (возможность наследования благоприобретенных признаков, т.е. признание передачи информации от фенотипа к генотипу. Название учения образовано от фамилии французского ученого Жана Батиста Ламарка; 2) жоффруизм (изменение организма под прямым воздействием среды, в особенности ранних зародышевых стадий). Учение носит имя французского натуралиста Этьена Жоффруа Сент-Илера; 3) дарвинизм (эволюция путем естественного отбора случайных изменений). Теорию сформулировал Чарлз Дарвин. 4) номогенез (развитие на основе закономерностей, а не случайностей). Автором концепции является Лев Семенович Берг. Основоположники этих учений в главном были правы, но в чем-то каждый из них ошибался, возможно, из-за признания абсолютной истины.
Теория Дарвина дала ответы на коренные вопросы биологии, в том числе и на то, почему морфология, физиология и поведение организмов соответствует условиям их обитания. Учение Дарвина имеет самое прямое отношение к медицине. Ведь медицина комплекс наук, изучающих человека не только больного, а и здорового, – существа, в основе своей, биологического, зависимого от природы и одновременно являющегося ее частью.
Эволюция кровеносной системы позвоночных животных
Между частями и органами тела любого организма в состоянии активной жизни постоянно происходит распределение веществ и газов. Транспорт жидкости и содержащихся в ней веществ может происходить на основе осмоса, за счет диффузии, молекул-переносчиков и специальных клеток. У высокоорганизованных организмов, в дополнение к этим процессам, появился эффективный способ перемещения жидкостей, осуществляемый механической работой специальных «насосов-сердец». У позвоночных имеется три обособленных полости, содержащих жидкости: система кровеносных сосудов, вторичная полость тела (целом) и межклеточные пространства, заполненные интерстициальной (тканевой) жидкостью. (Термин «жидкость» является не совсем корректным, поскольку это коллоидный раствор, но в научной и учебной литературе он закрепился давно, и заменить его другим, более адекватным термином довольно сложно). Отток жидкости из межклеточных пространств у позвоночных происходит через лимфатические сосуды, которые слепо начинаются в тканях и впадают в виде протоков в крупные кровеносные сосуды.
Важнейшей составной частью системы циркуляции является кровеносная система. Кровеносная система выполняет очень много функций. Она обеспечивает гомеостаз и целостность организма, осуществляя перенос кислорода питательных веществ ко всем органам, а углекислого газа к органам дыхания, доставляет гормоны к клеткам-мишеням, участвует в перераспределении тепла в организме. Наряду с транспортом веществ и газов, кровь выполняет защитную функцию в форме иммунных реакций. По кровеносным и лимфатическим сосудам происходит миграция клеток (в том числе раковых).
Кровеносная система имеется у различных типов бесхордовых (немертин, кольчатых червей, моллюсков, членистоногих, иглокожих) и хордовых животных. Хордовые – высший тип, подразделяющийся на три подтипа: бесчерепные, личиночнохордовые и черепные, или позвоночные. Кровеносная система хордовых сформировалась особым путем и независимо от состояния кровеносной системы бесхордовых. Её совершенствование происходило путем дифференциации (появление новых структур), интеграции (появление новых системных свойств) и интенсификации функций в ходе филогенеза (исторического развития) хордовых.
В медицинском аспекте представляет интерес эволюция кровеносной системы у бесчерепных и позвоночных животных, поскольку позволяет понять причину возникновения атавистических (aтавизм – возврат к предкам) пороков развития сердца и магистральных сосудов. Следует подчеркнуть, что кровеносная система исторически развивалась в тесной связи с другими системами (нервной, дыхательной, выделительной, пищеварительной) и рассмотрение ее, отдельно от других систем, обусловлено только соображениями дидактического характера (для облегчения восприятия учебного материала). В процессе эволюции усложнялись существующие органы и формировались новые (например, железы внутренней секреции), изменялась структура капилляров, появлялись новые регуляторные и структурные белковые молекулы. В крови человека насчитывается более трех тысяч различных белковых молекул.
Закладывается у эмбриона и сохраняется у взрослых особей в большей или меньшей степени.
Центральная нервная система, включая головной мозг, хорошо развита.
У различных групп организмов преимущественно развиваются различные органы чувств, достигая в развитии относительного совершенства.
Хорошо выражен внутренний скелет. Передний конец осевого скелета преобразован в череп.
Пищеварительный тракт расчленяется на ротовую полость, глотку, переднюю кишку (пищевод и желудок), среднюю кишку (тонкий кишечник), заднюю кишку (толстый кишечник).
Жаберные щели немногочисленные.
Получают развитие сложные органы дыхания.
и саморегуляции являются почки.
Замкнутая кровеносная система, имеется мускулистый орган для перекачивания крови – сердце.
Красные кровяные тельца – эритроциты содержат гемоглобин.
Органам кроветворения и депо крови является селезенка.
Развиты две пары конечностей.
Все представители подтипа ведут активный образ жизни. К этой группе относят: рыб, земноводных пресмыкающихся, птиц и млекопитающих.
Костные рыбы
Покровы тела – ктеноидная или циклоидная чешуя.
Боковая линия – воспринимает направление и силу тока воды.
Орган зрения – глаза имеют шаровидный хрусталик и плоскую роговицу, различают форму и цвет, веки отсутствуют.
Орган слуха – имеется только внутреннее ухо, среднее и наружное отсутствуют.
Обоняние – слепые мешки отходят от ноздрей, обоняние очень тонкое.
Размножение – раздельнополы, самка выметывает икру в воду, а самец покрывает ее семенем (молоками), т.е. оплодотворение наружное, имеются личиночные стадии.
Скелет – почти целиком костный, есть туловищный и хвостовой отделы, позвоночник образован костными позвонками с отростками, спинномозговой канал. Череп костный, состоит из мозговой коробки, челюстных костей, жаберных дуг и жаберных крышек.
Мышцы – сегментированы и представлены Z-образными туловищными мышцами, разделенными соединительно-тканными перегородками.
Нервная система – состоит из головного и спинного мозга и отходящих от них нервов. Головной мозг из пяти отделов. В поведении рыб проявляются безусловные и условные рефлексы.
Кровеносная система – замкнутая и состоит из двухкамерного сердца и сосудов. Один круг кровообращения.
– ротовая полость глотка пищевод желудок кишечник со слепыми отростками печень, желчный пузырь, поджелудочная железа, плавательный пузырь анальное отверстие.
Дыхательная система – четыре полные жабры и один ряд жаберных лепестков на внутренней стороне жаберной крышки.
Выделительная система – длинные лентовидные почки, по ним тянутся мочеточники, которые объединяются в непарный канал мочеполовой сосочек.
Земноводные
Покровы тела – кожа голая, участвует в дыхании, кожные железы постоянно выделяют увлажняющую слизь.
Боковая линия – у личинок и у хвостатых амфибий.
Орган зрения – глаза имеют веки, выпуклую роговицу, линзообразный хрусталик.
Орган слуха – внутреннее, среднее ухо с барабанной перепонкой, внутри слуховая косточка, стремечко.
Обоняние – наружные ноздри, ведущие в обонятельные капсулы.
Вкус – чувствительные клетки (во рту и по всему телу).
Размножение – раздельнополы, самки откладывают икру в воду, а самцы покрывают ее семенем, т.е. оплодотворение наружное, личинки напоминают мальков рыб.
Скелет – позвоночник, череп и скелет конечностей и их поясов. Грудная клетка отсутствует.
Мышцы – мощная и сложная мускулатура конечностей.
Нервная система – от отходит 10 пар нервов.
Кровеносная система – замкнутая, сердце трехкамерное, состоит из двух предсердий и одного желудочка. Два круга кровообращения.
Пищеварительная система – ротовое отверстие ротоглоточная полость пищевод, в полости рта язык желудок кишечник, поджелудочная железа, крупная печень, желчный пузырь, кишечник подразделен на тонкий и толстый, не переваренные остатки – через клоаку.
Дыхательная система – дышат легкими и кожей, в стадии зародыша жаберное дыхание.
Выделительная система – почки выводные протоки мочеточники открываются в клоаку.
Птицы
Покровы тела – тело покрыто перьями, на ногах роговые щитки.
Боковая линия – нет.
Орган зрения – глаза снабжены веками, в т.ч. и третьим веком – мигательная перепонка, обладают исключительной зоркостью, хорошо различают цвета.
Орган слуха – внутреннее, среднее и наружное ухо, хорошо слышат.
Обоняние – носовая полость, обоняние развито слабо.
Вкус – развиты слабо (чувствительные клетки во рту).
Размножение – раздельнополые. Оплодотворение внутреннее. Яйцекладущие. Яйца содержат желток и покрыты известковой скорлупой. Личиночных стадий нет.
Скелет – тонкие кости с обширными воздушными полостями придают легкость скелету, прочность за счет срастания ряда костей, позвоночник, череп, скелет конечностей и поясов конечностей.
Мышцы – грудные мышцы мощные, поднимают и опускают крылья, сложная мускулатура задних конечностей.
Нервная система – сложнее, чем у земноводных и пресмыкающихся. Масса равна массе спинного, полушария переднего мозга, связано с расширением двигательной активности и усложнением поведения. Мозжечок очень большой, это объясняется сложным движением птиц.
Кровеносная система – полное разделение большого и малого кругов кровообращения. Артериальная и кровь не смешиваются, сердце четырехкамерное, большое по объему, состоит из двух предсердий и двух желудочков, сердце работает интенсивно.
Пищеварительная система – ротовая полость, язык, слюнные железы, длинный пищевод с зобом, тонкостенный двухкамерный желудок, железистный отдели тостостенный мускульный отдел. Кишечник тонкий и толстый. В тонком лежит поджелудочная железа, толстый укорочен и заканчивается клоакой.
Дыхательная система – легочное дыхание, трахея разделяется на два , нижняя с голосовыми связками. Легкие имеют зубчатое строение, воздушные мешки.
Выделительная система – почки мочеточники клоака. Мочевого пузыря нет.
Млекопитающие
Покровы тела – тело покрыто шерстью – волосами, среди которых имеются осязательные волоски – вибриссы; два типа кожных желез: сальные и потовые.
Боковая линия – нет.
Орган зрения – глаза снабжены веками, но третье веко рудиментарное, острота зрения зависит от условий жизни. Цветовое зрение слабее, чем у птиц, либо отсутствует.
Орган слуха – имеют внутреннее, среднее и наружное ухо, слух обычно хорошо развит, у некоторых есть звуковая локация.
Обоняние – носовая полость с носовыми раковинами, покрыты эпителием, обоняние тонкое.
Вкус – почки, чувствительные клетки, которые в виде сосочков погружены в поверхность языка.
Размножение – раздельнополые. Оплодотворение внутреннее. Зародыш, как правило, развивается в организме самки, живородящие.
Скелет – совершенный скелет, подотделы: туловище и голова (череп), скелет конечностей и поясов конечностей. У позвонков плоские поверхности между которыми расположены хрящевые диски. Крупная мозговая коробка.
Мышцы – дифференцирована и представлена множеством разнообразно расположенных мускулов, характерно наличие диафрагмы, развита подкожная мускулатура, мимическая на лице.
Нервная система – головной мозг характеризуется крупными размерами. Разрастается кора головного мозга. У большинства видов она не гладкая, а образует борозды, увеличивающие ее площадь. Крупный мозжечок дифференцирован на отделы.
Кровеносная система – сложно устроена. Два круга кровообращения, совершенные сосуды, аорта, капилляры.
Пищеварительная система – сложная, пищеварительный тракт длинный. Предротовая полость (зубы, щеки, челюсти), ротовая полость, зубы, язык глотка пищевод желудок с железами кишечник толстый и тонкий. Система желез: печень, желчный пузырь, поджелудочная железа. Заканчивается анальным отверстием.
Дыхательная система – легкие, трахея и бронхи, которые хорошо развиты. В области легких бронхи делятся на большое число веточек. Самые мелкие заканчиваются альвеолами – пузырьками, в которых происходит газообмен.
Выделительная система – почки, состоящие из наружного (коркового) и внутреннего (мозгового) слоев мочеточники мочевой пузырь мочеиспускательный канал. Частично выделительную функцию выполняют потовые железы.
1.1. Происхождение и функции покровов тела.
Эпидермис покровов тела образуется из эктодермы, а соединительная ткань кожи – из мезодермы.
Покровы тела выполняют следующие функции:
Защита от вредных воздействий среды;
Восприятие раздражений;
Участие в обмене веществ;
Покровы тела – дополнительный орган дыхания и выделения, а у членистоногих это наружный скелет.
1.2. Эволюция покровов тела у беспозвоночных животных.
Покровы тела у беспозвоночных животных образованы преимущественно эктодермой и ее производными. Так, кожно-мускульный мешок плоских червей состоит из покровной ткани – тегумента, который представляет собой многоядерную неклеточную структуру и трех слоев мышц – продольных, поперечных и дорзовентральных. Движения, осуществляемые ими, медленны и несовершенны. У турбеллярий тело покрыто ресничками, участвующими в движении.
Кожно-мускульный мешок круглых червей состоит из многослойной, эластичной и малорастяжимой кутикулы, гиподермы, представляющей собой единую цитоплазматическую массу, не разделенную на отдельные клетки и содержащую большое количество ядер, и одного слоя продольных и гладких мышц. Кутикула выполняет в основном защитную функцию. Мышцы располагаются в виде двух продольных тяжей – на брюшной и спинной сторонах тела. Их поочередное сокращения обеспечивает энергичные сгибательные и разгибательные движения и быстрое перемещение тела в пространстве.
Тело кольчатых червей покрыто кожно-мускульным мешком, состоящим из одного слоя эпителия и двух слоев мышц: наружного слоя кольцевых и внутреннего слоя, образованного продольными мышечными волокнами.
Таким образом, эволюция покровов тела у беспозвоночных шла от мерцательного эпителия турбеллярий, т.е. ресничных червей, выполняющих у них функцию примитивных органов движения, к плоскому, лишенному мерцательных выростов эпителию сосальщиков, ленточных, круглых и кольчатых червей.
Покровы тела членистоногих представлены хитинизированной кутикулой, защищающей органы от внешних вредных воздействий и представляющая собой наружный скелет, к которому прикрепляются обособленные мышцы, состоящие из поперечнополосатых мышечных волокон.
1.3. Эволюция покровов тела у хордовых животных.
У хордовых животных в коже присутствует два слоя: верхний – эктодермальный эпидермис и нижний – мезодермальный кориум или собственно кожа, или дерма.
Эволюция кожных покровов у хордовых шла по пути перехода от однослойного эпителия к многослойному, а также в изменении соотношения между слоями в сторону большего развития собственно кожи.
Для низших хордовых, в частности для бесчерепных, характерна слабая степень дифференцировки обоих слоев кожи. Эпидермис однослойный цилиндрический, ограниченный снаружи тонкой пористой кутикулой, содержащей одноклеточные слизистые железы. Под эпидермисом лежит тонкая опорная пластинка, а под ней слабовыраженный слой студенистой соединительной ткани, представляющей собственно кожу или дерму.
У высших хордовых животных, т.е. у позвоночных, покровы тела состоят не из однослойного, а из многослойного эктодермального эпидермиса и мезодермального волокнистого кориума, или собственно кожи. Эпидермис имеет два слоя. Клетки нижнего, так называемого камбиального, слоя служат для воспроизведения новых слоев клеток, а клетки верхнего слоя имеют защитную функцию. Верхний слой имеет многочисленные слизистые и другие железы, в нем у большинства наземных животных за счет ороговевающих клеток образуются роговые щитки, чешуйки, перья, волосы, ногти, когти, рога, копыта и т.д. В верхнем слое эпидермиса клетки дифференцируются, гибнут и слущиваются.
Глава 2.
Эволюция скелета.
2.1. Происхождение и функции скелета животных.
Опорные образования у беспозвоночных животных, представляющие им постоянную форму тела очень разнообразны. Они имеют экто-, энто- и мезодермальное происхождение. У позвоночных животных скелет имеет в основном мезодермальное происхождение.
Скелет в организме животного выполняет разнообразные функции:
Обеспечение постоянной формы тела;
Пассивная часть опорно-двигательного аппарата;
Защита от механических и других воздействий;
Кроветворная функция.
2.2. Эволюция скелета в ряду беспозвоночных животных.
У губок опорные образования представлены иглами, имеющих различный химический состав.
У кишечнополостных появляется плотная опорная пластинка (мезоглея), которая занимает место между экто- и энтодермой. Скелет коралловых полипов развивается из эктодермы. У членистоногих наружный скелет представлен хитинизированным покровом, включающим функции защиты от механических повреждений и наружного скелета, к которому прикрепляются впервые появившиеся у членистоногих поперечнополосатые мышцы.
Двустворчатые и брюхоногие моллюски имеют раковины, образованные выделениями мантии. Головоногие моллюски имеют сложные хрящевые образования, защищающие нервные центры и органы чувств.
2.3. Эволюция скелета у хордовых животных.
Как и у беспозвоночных, скелет хордовых животных выполняет функцию защиты органов и служит опорой для органов передвижения.
Осевой скелет в процессе эволюции претерпел большие изменения.
У низших хордовых осевым скелетом является хорда, а у высших она постепенно заменяется развивающимися позвонками. В позвонках различают тело, верхние и нижние дуги.
Так у круглоротых хорда сохраняется в течении всей жизни, но появляются закладки позвонков, представляющие собой небольшие хрящевые образования, метамерно располагающиеся над хордой. Они называются верхними дугами.
У примитивных рыб кроме верхних дуг появляются нижние дуги, а у высших рыб и тела позвонков. Тела позвонков у большинства рыб и вышестоящих животных формируется из ткани, окружающей хорду, а также из оснований дуг. С телами позвонков срастаются верхние и нижние дуги. Концы верхних дуг срастаются между собой, образуя спинномозговой канал. На нижних дугах появляются отростки, к которым прикрепляются ребра. У рыб два отдела позвоночника – туловищный и хвостовой. Остатки хорды у рыб сохраняются между телами позвонков.
У земноводных на ранних стадиях развития хорда заменяется позвоночником. В позвоночнике уже четыре отдела: шейный, грудной, крестцовый и хвостовой. В шейном отделе всего один позвонок, грудной состоит из пяти позвонков. К грудным позвонкам прикрепляются небольшие ребра, заканчивающиеся свободно. Крестцовый отдел, как и шейный, включает один позвонок, который является опорой для костей таза и задних конечностей. Хвостовой отдел у бесхвостых амфибий срастается в одну кость, а у хвостатых состоит из большого числа позвонков.
У рептилий пять отделов в позвоночнике: шейный, грудной, поясничный крестцовый и хвостовой. В шейном отделе у различных видов пресмыкающихся разное число позвонков, но самое большое восемь. Первый позвонок называется атлантом и имеет форму кольца, а второй – эпистрофеем и имеет зубовидный отросток, на котором вращается первый позвонок. В грудном отделе число позвонков не постоянно, к ним прикрепляются ребра, большинство из которых соединяется с грудиной, впервые образуя у высших животных грудную клетку. В грудопоясничном отделе всего 22 позвонка, а в крестцовом два. К позвонкам поясничного и крестцового отделов также прикрепляются ребра. В хвостовом отделе число позвонков варьирует, иногда их несколько десятков.
У птиц позвоночный столб сходен с позвоночником рептилий, но обладает определенной специализацией в связи хвостовой. а позвоночника - туловищный ются ребра. у ы уется из ЫВДр, окружающей хорду, а также из основания дуг.образования, с образом жизни. Шейный отдел включает до 25-х позвонков, что обеспечивает большую подвижность.
У млекопитающих в позвоночнике пять отделов: шейный грудной, поясничный, крестцовый и хвостовой. В шейном отделе семь позвонков, в грудном непостоянное число позвонков (от 9 до 24, но чаще 12-13). К грудным позвонкам присоединяются ребра, большие количество из которых соединяется с грудиной. Поясничный отдел включает от трех до девяти позвонков. Крестцовые позвонки срослись, образовав крестец, а хвостовой отдел позвоночника включает разное количество позвонков у различных видов млекопитающих.
Скелет головы. Скелетом головы является череп. Он расположен на переднем конце скелета и состоит из двух частей: черепной коробки и висцерального скелета, которые различаются между собой как по происхождению, так и по выполняемой функции. Черепная коробка служит вместилищем головного мозга, а висцеральный скелет дает опору органам передней части пищеварительного канала.
В процессе эволюции наибольшие изменения происходят в висцеральном отделе. У зародышей всех позвоночных, а у низших позвоночных в течении всей жизни, висцеральный скелет состоит из дуг, охватывающих переднюю часть пищеварительной трубки. У рыб они дифференцируются на челюстную дугу (захват пищи), подъязычную (прикрепление к черепной коробке) и жаберную дугу (прикрепления жабр).
У наземных животных висцеральный скелет сильно преобразуется и редуцируется: верхняя часть челюстной дуги срастается с дном черепной коробки, из подъязычной дуги образуются небольшие косточки, входящие в состав среднего уха. Вторая и третья жаберные дуги у млекопитающих образуют щитовидный хрящ, а из четвертой и пятой дуг формируются остальные хрящи гортани.
Скелет конечностей. Существует два типа свободных конечностей. Это плавники рыб и пятипалые конечности млекопитающих животных. Пятипалые конечности позвоночных животных имеют очень разнообразное строение, что связано с выполнением ими различных функций. Например, роющие конечности крота, плавающие – у тюленя, лазающие – у обезьян и т.д. Но тем не менее, несмотря на различие, конечности позвоночных животных сохраняют общий план строения, что доказывает общность их происхождения.
Впервые конечности появились у рыб и были представлены плавниками. Это парные грудные и брюшные плавники. У большинства рыб плавники состоят из радиальных тонких костных лучей и выполняют функцию изменения направления плавания, а не опоры тела. У кистеперых рыб намечается укрупнения лучей за счет срастания и использования плавников в качестве опоры и передвижения по твердому основанию пересыхающих водоемов. Поэтому плавники древних кистеперых рыб явились основой для развития конечностей позвоночных животных. Важной чертой преобразования плавников в конечности наземных позвоночных явилась замена прочного соединения скелетных элементов подвижным соединением в виде суставов. В результате этого конечность превратилась в сложный подвижный рычаг, в котором выделяют три кости: плечо, предплечье и кисть. Имеется два пояса конечностей – плечевой и тазовый.
Далее эволюция передней конечности шла по пути удлинения плеча и предплечья, укорочения запястья, уменьшения количества костей в запястном отделе (у амфибий – 3 ряда, у млекопитающих – 2 ряда) и удлинения дистальных отделов, т.е. фаланг пальцев.
Скелет руки человека также характеризуется общим планом строения с передними конечностями позвоночных животных, но наряду с этим имеет и важные отличия, так как руки человека являются не только оружием труда, но и ее результатом и способна выполнять многообразные действия.
2.4. Аномалии и пороки развития скелета у человека.
1. Наличие ребер у нижнего шейного или у первых поясничных позвонков . В соответствии с эволюцией позвоночных у человека во время эмбрионального развития во всех отделах позвоночника закладываются ребра, но впоследствии они сохраняются только в грудном отделе, а в других отделах ребра редуцируются. Но иногда у человека наблюдаются подобные атавизмы.
2. Наличие хвостовых позвонков . Во время эмбриогенеза у человека, как и позвоночных животных, закладывается 8-11 хвостовых позвонков, затем они редуцируются и остается 4-5 недоразвитых позвонков, образуя копчик. Иногда проявляются атавистические признаки в виде присутствия хвостового отдела позвоночника.
3. Spina bifida – это нередкая аномалия, которая возникает при нарушении срастания верхних дуг позвонков. Она чаще проявляется в пояснично-крестцовой области позвоночника и в зависимости от глубины и протяженности расщепления может иметь разную степень тяжести.
4. Наличие в барабанной полости только одной слуховой косточки - столбика . Это нарушение, соответствующее строению звукопередающего аппарата земноводных и пресмыкающихся, является результатом неправильной дифференцировки элементов челюстной жаберной дуги в слуховые косточки. Это рекапитуляция основных этапов филогенеза висцерального черепа в онтогенезе.
5. Гетеротопия пояса верхних конечностей . Это перемещение пояса верхних конечностей из шейного отдела на уровень 1-2 грудного позвонков. Данная аномалия называется болезнью Шпренгеля или врожденным высоким стоянием лопатки. Она выражается в том, что плечевой пояс с одной или с двух сторон находится выше нормального положения на несколько сантиметров. Механизм такого нарушения связан как с нарушением перемещения органов, так и с нарушением морфогенетических корреляций.
6. Полидактилия – результат развития закладок дополнительных пальцев, характерных для далеких предковых форм.
7. Плоскостопие, косолапость, узкая грудная клетка, отсутствие подбородочного выступа – атавистические аномалии скелета, которые часто встречаются и являются аноболиями (надставки), возникшими в ходе филогенеза приматов.
Ветуликолии впервые найденные в Гренландии в 1911 году, попадаются в кембрийских отложениях разных регионов мира – от Китая до Канады. Слепые, но обладавшие широким ртом, они могли плавать благодаря движениям хвоста. С экологической точки зрения ветуликолии напоминали миниатюрных китовых акул, отфильтровывающих планктон и органическую взвесь из толщи воды.
На сегодняшний день науке известно 14 видов этих животных, но из-за крайне неудовлетворительной сохранности их родственные связи, а также подробности морфологии и внешнего вида до сих пор оставались плохо изученными. Известно лишь, что тело Vetulicolia четко делилось на два отдела – полый передний и сегментированный задний. Строение переднего отдела говорит о том, что ближайшими родственниками ветуликолий могли быть примитивные хордовые вроде сальпов и асцидий, а хвост навевает мысли о членистоногих. Разобраться в родственных связях группы помогла новая находка.
Недавно группа австралийских палеонтологов под руководством Диего Гарсия-Беллидо из университета Аделаиды обнаружила на острове Кенгуру окаменелости нового вида ветуликолий. Ученые назвали его Nesonektris aldridgei, в честь известного исследователя этой группы Дика Элдриджа из британского университета Лестера. Родовое имя животного в переводе с греческого языка означает «островной пловец».
Незонектрисы вырастали примерно до 13 см в длину. Любопытной особенностью их сохранности является то, что хвосты и передние отделы часто встречаются по отдельности, то есть были соединены непрочно и оказывались раздроблены вскоре после смерти животных. Еще больше заинтересовала палеонтологов трубка, проходящая вдоль тела наподобие кишки: она была разделена перегородками на отдельные блоки.
«Это совершенно несовместимо с версией о кишечнике (который представляет собой полую трубку), но хорошо соответствует хрящевой хорде (или нотохорду), – отметил Гарсия-Беллидо. – Таким образом, теперь мы можем делать выводы о том, где именно эта группа располагалась на древе жизни».
Нотохорд встречается практически у всех позвоночных на эмбриональных этапах развития, по мере взросления обычно уступая место позвоночнику. Некоторые примитивные хордовые сохраняют нотохорд на протяжении всей жизни, а у других, вроде асцидий, он имеется только на ранних стадиях развития. Если у ветуликолий действительно имелась хорда, то это помещает их в группу предков или как минимум «двоюродных братьев» всех остальных хордовых, и в том числе человека.
«Они являются близкими родственниками позвоночных. У Vetulicolia имеется длинный хвост, поддерживаемый жестким стержнем, напоминающим хорду, которая является предшественником позвоночника и служит уникальным признаком позвоночных и их родственников», – сказал австралийский палеонтолог. Теперь Гарсия-Беллидо и его команда намерены переизучить известные остатки других ветуликолий, чтобы попытаться найти нотохорд и у них. Кроме того, будут продолжены раскопки на острове Кенгуру, где благодаря редкому стечению обстоятельств сохранились в ископаемом состоянии отпечатки мягких тканей животных кембрийского периода.
Статья A new vetulicolian from Australia and its bearing on the chordate affinities of an enigmatic Cambrian group опубликована в журнале BMC Evolutionary Biology
Челюсти позвоночных животных, как считается, произошли из передней пары жаберных дуг их древнейших рыбообразных предков. Американские ученые наконец-то обнаружили документальное подтверждение этой гипотезы в окаменелости кембрийского организма Metaspriggina.

Metaspriggina walcotti. Реконструкция: Marianne Collins
Согласно современным научным представлениям, челюсти и несколько мелких косточек внутреннего уха человека ведут свою родословную от жаберных дуг – своеобразных скелетных структур, в кембрийское или даже докембрийское время поддерживавших жабры примитивных рыбообразных позвоночных. Однако в связи с плохой сохранностью и неполнотой геологической летописи обнаружить эти дуги на месте их расположения в теле организма до последнего времени не представлялось возможным.
Только что профессор Саймон Моррис из британского Кембриджского университета и его коллега, доктор Жан-Бернар Карон из университета Торонто и Королевского музея Онтарио описали жаберные структуры раннего хордового Metaspriggina walcotti, жившего 505 млн лет назад. Окаменелости этой метасприггины уникальной сохранности были найдены в 2012 году в знаменитом местонахождении раннекембрийской фауны Берджес Шейл вместе с четырьмя десятками других фоссилий.
«Детали этих окаменелостей ошеломляют, – заявил Моррис. – Даже глаза прекрасно сохранились и отлично видны». Два крупных глаза метасприггины оказались вынесены на самый конец морды и оттого кажутся выпученными. А неподалеку от них ученые разглядели слегка изогнутые палочки – те самые жаберные дуги, которым суждено было со временем дать начало челюстям и слуховым костям позвоночных.
«Эти дуги, как уже давно известно, играли ключевую роль в эволюции позвоночных, в том числе в возникновении у них челюстей и крошечных косточек, занятых передачей звука у млекопитающих, – отметил британский профессор. – До сих пор, однако, нехватка качественных окаменелостей приводила к тому, что наши представления о расположении этих арок у первых позвоночных были чисто гипотетическими».
Оказалось, что жаберные дуги располагались в теле животного парами, что как нельзя лучше подтверждает гипотезу происхождения от них пары челюстей. При этом первая пара дуг у Metaspriggina была еще и толще всех остальных, что, возможно, является первым эволюционным шагом на пути к возникновению челюстноротых.
Дополнительно проанализировав строение и расположение мышц метасприггины, палеонтологи пришли к выводу, что это животное было активным и подвижным пловцом, не уступающим современной форели. А пара крупных глаз и обонятельный орган позволяли им отлично ориентироваться в море кембрийского периода.

Хордовые имеют возраст около 700 млн. лет назад, но хорошая геологическая история их эволюции начинается только в Кембрии. В океанах Кембрия и Ордовика известны только такие хордовые животные, какие не имели челюстей.
В Силуре получилось, случилось, произошло сформировались амфибии имевшие челюсти… От них происходят акулы и скаты, осетры овые рыбы, костистые рыбы и амфибии.
В Каменноугольном периоде появились наземные животные, — анопсиды… От анопсид происходят синопсиды и диопсиды. Эта классификация как бы учитывает количество височных ям, но реально это не так. Например, к диопсидам относят анопсидных анкилозавров, и птиц — имеющих открытые високчные углубления впадиныямы… К синопсидам относят однопроходных, по строению черепа близких к динозаврам и к птицам. Однопроходных относят к млекопитающием из-за имеющихся у них кожаных выделений и шерсти. Но выделения однопроходных подобны не молоку, а секрету копчиковой железы птиц, а шерсть такая же, как шерсть птерозавров…
В общем, от анопсид развилось несколько классов животных, по-разному сочетающих и развивающих их признаки. Эти стволы: ящерицы и змеи, крокодилы, динозавры (включая однопроходных), плезиозавры, тероморфы (включая ихтиозавров).
Потомки тероморфов, млекопитающие, известны еще в Триасе и Меле. Но возможность широко распространиться, млекопитающие животные получили только в эпоху Палеогена и Неогеновую эпоху. Это стало возможным после массового вымирания в Верхнем Меле множества видов во всех классах позвоночных.
В Палеогене появились млекопитающие со складчатой корой головного мозга: хищные, копытно-дамановые, обезьяны. Предком всех этих отрядов млекопитающих были какие-то лемуры. На лемуров похожи все древние млекопитающие, имеющие складчатую кору головного мозга.
Акулы потеряли все свои кости в процессе эволюции
29.05.2015 14:22
Ископаемая акула, найденная в западной Австралии, буквально перевернула представления ученых об эволюции этой группы рыб. Если еще недавно опасных морских хищниц считали достаточно примитивными созданиями, то теперь биологам придется относиться к ним, как к весьма продвинутым существам, проделавшим длинный эволюционный путь.

Gogoselachus lynnbeazleyae. Реконструкция: John Long
Революцию в ихтиологии устроил палеонтолог университета Флиндерса Джон Лонг. Три десятилетия Лонг раскапывает отложения девонской формации Гого (Gogo formation) в регионе Кимберли. В 2005 году он нашел там окаменевший скелет акулы, жившей в теплом тропическом море 380 млн лет назад. Изучение и описание животного затянулось, и статья о Gogoselachus lynnbeazleyae увидела свет лишь сейчас.
«Акул принято считать примитивными рыбами на том основании, что их скелет состоит из хряща, а кости у них никогда не образуются, – рассказывает профессор Лонг. – Однако теперь мы переворачиваем эту идею с ног на голову, утверждая, что ранние ископаемые акулы на самом деле имели настоящий костный скелет, и лишь впоследствии его потеряли».
Действительно, костную ткань у современных акул можно найти лишь в корнях зубов, а скелет и даже череп состоят из хрящевой ткани, считающейся предшественницей ткани костной. Эти представления распространяли и на всех акул прошлого. Но когда Лонг взглянул на хрящ гогоселяхуса под сильным увеличением с помощью микротомографии, то увидел в нем настоящие остеоциты – клетки, из которых состоят кости.
«Наша ископаемая акула впервые показала настоящую костную конструкцию, связывающую воедино крошечные хрящики. То есть мы видим акулу, которая на самом деле произошла от кого-то, у кого в скелете было намного больше костей. А на другом конце этой линии располагаются современные акулы, полностью утратившие кости и ставшие хрящевыми. Таким образом, наше ископаемое позволяет наблюдать эволюцию тканей, и объясняет причины, по которым современные акулы стали настолько успешными в наши дни – они просто отказались от костей, чтобы стать более легкими», – пояснил Лонг.
«Это действительно интересное открытие, – отметил профессор палеонтологии Упсальского университета Пер Альберг. – Скелеты современных акул состоят из своеобразной ткани, называемой призматическим кальцинированным хрящом. Этот хрящ минерализован и выглядит не как твердые листы, а как мозаика из крошечных минеральных призм. Такая ткань довольно сильно отличается от кости, и ее происхождение пока не очень хорошо понятно. Новая акула из Гого показывает, что, кажется, ранняя версия призматического кальцинированного хряща, в отличие от современного, имела между призмами зазоры, заполненные костными клетками».
«Изучение акул очень похоже на разгадывание грандиозной головоломки, – добавил профессор Лонг. – Они появились за 250 млн лет до последних динозавров и не сильно изменились с тех пор, удачно попав в выигрышную формулу. Но хотя их внешность осталась почти прежней, строение тканей претерпело серьезные изменения».
Выяснить все эти подробности Лонгу удалось благодаря специфическим условиям сохранности ископаемого материала в формации Гого. Обычно девонские рыбы сохраняются в окаменелостях сильно деформированными, расплющенными давлением осадочных пород. Но здесь, в Кимберли, древние рыбы дошли до нас трехмерными и объемными благодаря карбонатным конкрециям, образовавшимся на месте губко-водорослевого рифа.
«В те времена тут кипела жизнь, обитали многие виды рыб, например, давно вымершие бронированные плакодермы и ранние костные рыбы, потомки которых доминируют сегодня. Мы надеялись встретить тут много акул, но по какой-то причине они не были распространены на данном рифе», – добавил Лонг.
По этой причине находка скелета 75-сантиметровой акулы привлекла большое внимание. В руки ученых попали обе ветви нижней челюсти, фрагменты плечевого пояса, поддерживавшего грудные плавники, жаберные арки, около 80 зубов и несколько сотен чешуй. Образец был подвергнут множеству самых разных исследований, и одно из них привело к сенсационному результату, заставляющему пересмотреть как раннюю эволюцию акул, так и отношение к ним как к примитивной, задержавшейся в своем развитии группе.
Статья First Shark from the Late Devonian (Frasnian) Gogo Formation, Western Australia Sheds New Light on the Development of Tessellated Calcified Cartilage опубликована порталом PLOS ONE
Doi: 10.1371/journal.pone.0126066
Согласно новым данным американских ученых, птицы не являются потомками динозавров и происходят от особой группы архозавров, обособившейся от гигантских ящеров в далеком прошлом.

Scansoriopteryx. Реконструкция: Matt Martyniuk
Сенсационное открытие, способное буквально перевернуть с ног на голову всю современную палеонтологию, сделали Стивен Черкас из Музея динозавров в городе Блэндинг и Алан Федуччия из университета Северной Каролины. С помощью своей новой методики они изучили остатки крохотного пернатого ящера Scansoriopteryx и пришли к выводу о том, что нет никаких оснований считать его динозавром.
Scansoriopteryx, название которого переводится на русский язык примерно как «крылолаз» или «лазокрыл», был мелким, примерно с воробья, архозавром. Остатки единственной, неполовозрелой особи скансориоптерикса нашли в начале 21 века в юрских отложениях китайской провинции Ляонин. Судя по дошедшим до наших дней ископаемым, Scansoriopteryx был устроен примитивнее знаменитого археоптерикса и хорошо умел лазать по деревьям, планируя с них с помощью небольших крыльев.
До последнего времени скансориоптерикса относили к целурозаврам – группе теропод, от которых, по мнению большинства ученых, и произошли современные птицы. Однако исследование Черкаса (который, кстати, и открыл Scansoriopteryx) и Федуччия заставляет пересмотреть этот подход. Дуэт ученых использовал трехмерную микроскопию и фотографирование при низких углах освещения, чтобы прояснить структуры, нечетко рассмотренные прежде. Благодаря этому палеонтологам удалось уточнить естественные очертания костей таза, хвоста и конечностей, а заодно и обнаружить удлиненные сухожилия, тянувшиеся вдоль хвостовых позвонков, как у Velociraptor.
Тем не менее, большинство полученных данных свидетельствуют о том, что скансориоптериксу не хватало фундаментальных структурных особенностей скелета, чтобы быть причисленным к динозаврам. Скорее, он является потомком ранних архозавров, освоивших лазание по деревьям задолго до появления ужасных ящеров Соответственно, и птицы, у самых корней родословного древа которых располагается Scansoriopteryx, являются не потомками динозавров, а в лучшем случае их двоюродными племянниками.
Между тем у самого скансориоптерикса отчетливо видны типично птичьи адаптации вроде удлиненных передних конечностей, превратившихся в оперенные крылья, специализированной полулунной кости запястья и приспособленных к усаживанию на ветки лап. Скорее всего, это животное находилось в начале освоения полета, в который отправлялось, планируя с ветвей деревьев.