Они исполняют роль пограничного барьера, отделяя ниже лежащие ткани от окружающей среды. Первичные покровы растения состоят только из живых клеток. Вторичные и третичные покровы – в основном из мертвых с толстыми клеточными стенками.
Основные функции покровных тканей:
· защита растения от высыхания;
· защита от попадания вредных микроорганизмов;
· защита от солнечных ожогов;
· защита от механических повреждений;
· регуляция обмена веществ между растением и окружающей средой;
· восприятие раздражения.
Первичная покровная ткань – эпидерма, эпидермис. Состоит из живых клеток. Образуется из апикальных меристем. Покрывает молодые растущие стебли и листья.
Эпидерма сформировалась у растений в связи с выходом из водной среды обитания на сушу с целью предотвращения от высыхания. Кроме устьиц, все клетки эпидермы плотно соединены между собой. Наружные стенки основных клеток толще остальных. Вся поверхность покрыта слоем кутина и растительных восков. Этот слой называется кутикулой (кожица). Она отсутствует на растущих корнях и подводных частях растений. При пересыхании проницаемость кутикулы значительно ослабляется.
Кроме основных клеток, в эпидермисе имеются и другие, в частности волоски, или трихомы. Они бывают одноклеточными и многоклеточными (рис.2). Функционально они увеличивают поверхность эпидермы, например, в зоне роста корня, служить механической защитой, цепляться за опору, уменьшать потери воды. Ряд растений имеют железистые волоски, например, крапива.
Только у высших растений в эпидермисе имеются устьица, которые регулируют обмен воды и газов. Если кутикулы нет, то и отсутствует потребность в устьицах. Устьица – это группа клеток, образующих устьичный аппарат, который состоит из двух замыкающих клеток и примыкающих к ним клеток эпидермы – побочных клеток. Они отличаются от основных эпидермальных клеток (рис.3
). Замыкающие клетки отличаются от окружающих их клеток формой и присутствием большого количества хлоропластов и неравномерно утолщенными стенками. Те, которые обращены друг к другу, толще остальных (рис.4)
Между замыкающими клетками образуется устьичная щель, которая ведет в подустьичное пространство, называемое подустьичной полостью. Замыкающие клетки обладают высокой фотосинтетической активностью. В них содержится большое количество запасного крахмала и многочисленные митохондрии.
Число и распределение устьиц, типы устьичных аппаратов широко варьирует у различных растений. Устьица у современных мохообразных отсутствуют. Фотосинтез у них осуществляет гаметофитное поколение, а спорофиты к самостоятельному существованию не способны.
Обычно устьица располагаются на нижней стороне листа. У плавающих на водной поверхности растений – на верхней поверхноси. У листьев злаков устьица часто располагаются равномерно с обеих сторон. Такие листья освещаются сравнительно равномерно. На 1мм2 поверхности может располагаться от 100 до 700 устьиц.
Вторичная покровная ткань (перидерма). Эта ткань приходит на смену эпидерме, когда зеленый цвет однолетних побегов сменяется коричневым. Она многослойна и состоит из центрального слоя камбиальных клеток - феллогена. Клетки феллогена, делясь, наружу откладывают слой феллемы, а внутрь – феллодерму (рис.5).
Феллема, или пробка. Сначала состоит из живых тонкостенных клеток. Со временем их стенки пропитываются суберином и растительными восками и отмирают. Содержимое клетки наполняется воздухом.
Функции феллемы:
· предотвращает потерю влаги;
· защищает растение от механических повреждений;
· защищает от болезнетворных микроорганизмов;
· обеспечивает термоизоляцию, так как клетки заполнены воздухом.
Клетки феллогена, расположенного в самой эпидерме, подлежащем субэпидермальном слое, реже – в глубоких слоях первичной коры, являются генерирующей основой первичной коры.
Статьи и публикации:
Историческая справка
Первые работы по выделению и изучению белковых препаратов были выполнены еще в 18 в., однако в тот период исследования белков носили описательный характер. В начале 19 в. были сделаны первые анализы элементного состава белков (Ж.Л. Гей-Лю...
Микробиологический контроль производства кисломолочных продуктов
Задачи микробиологического контроля сводятся к обеспечению надлежащей направленности микробиологических процессов и соблюдению санитарно-гигиенических условий производства.
Исходя из этого, санитарно-микробиологический контроль, производ...
Учение Павлова о двух сигнальных системах действительности.
Возрастные особенности развития и взаимодействия первой и второй сигнальной
системы. Роль функциональной асимметрии мозга в развитии речи
Сигнальные системы – это системы нервных процессов, временных связей и реакций, формирующиеся в головном мозге в результате воздействия внешних и внутренних раздражений и обеспечивающие тонкое приспособление организма к окружающей среде. ...
Снаружи органы растения покрыты тканями , называемыми покровными . Они защищают внутренние части растения от неблагоприятных внешних воздействий, регулируют газообмен и транспирацию (испарение воды).
Различают три типа покровных тканей — эпидерму , перидерму и корку .
Эпидерма
Эпидерма покрывает листья, травянистые стебли, части цветков и многие плоды. Она чаще всего состоит из одного слоя живых клеток, оболочки которых обычно извилистые, благодаря чему они прочно соединяются между собой. Наружные, граничащие с внешней средой оболочки клеток более толстые и покрыты слоем кутикулы, воска, а также у многих растений волосками, чешуйками, усиливающими защитную функцию эпидермы. Форма клеток эпидермы сильно варьирует и зависит от формы органа, который она покрывает. Так, на листьях с широкой листовой пластинкой эпидермальные клетки также широкие, а на листьях линейных и на стеблях они сильно вытянуты (рис. 9).
Большую часть объёма клетки эпидермы занимает вакуоль, заполненная клеточным соком. Цитоплазма тонким слоем прилегает к оболочке клетки. В ней присутствуют лейкопласты, иногда хлоропласты (у водных и теневыносливых растений). Среди основных клеток эпидермы располагаются устьица , через которые происходит газообмен и транспирация. Они состоят из двух замыкающих клеток бобовидной формы, между которыми имеется щель, сообщающаяся с межклетниками ассимиляционной ткани. В замыкающих клетках устьиц содержатся хлоропласты.
Замыкающие клетки вместе с окружающими их клетками эпидермы, называемыми побочными, образуют устьичный аппарат. Устьица обладают способностью автоматически открываться и закрываться по мере надобности растения. Управляет этим процессом тургор замыкающих клеток. Дело в том, что стенки их со стороны устьичной щели сильно утолщены, а с противоположной — тонкие. Днем, когда в клетках идёт процесс фотосинтеза, концентрация клеточного сока повышается, вода из прилегающих клеток эпидермы поступает в замыкающие клетки и тургор в них возрастает, в результате этого тонкие стенки их растягиваются и тянут за собой утолщенные, которые открывают щель. Ночью концентрация сахара в замыкающих клетках снижается, тургор падает, и устьица закрываются. У большинства растений устьица располагаются на нижней стороне листа, но у листьев, расположенных вертикально (у злаков), присутствуют на обеих сторонах, а у листьев, плавающих на поверхности воды, устьица располагаются на верхней стороне листа.
Перидерма
Перидерма — это вторичная покровная ткань (рис. 10), которая развивается взамен эпидермы на многолетних стеблях и корнях из пробкового камбия, или феллогена. Феллоген возникает из клеток эпидермы или основной паренхимы, расположенных под эпидермой или в более глубоких слоях коры. Клетки его делятся параллельно поверхности органа, откладывая кнаружи правильные радиальные слои клеток, превращающиеся в слои пробки. Стенки клеток пробки пропитываются суберином и воском, утолщаются и становятся непроницаемыми для воды, что влечёт за собой отмирание протопластов. Материал с сайта
Внутрь от себя феллоген откладывает в небольшом количестве клетки, которые остаются живыми, хлорофиллоносными; они образуют ткань, называемую феллодермой. Таким образом, перидерма — это сложная многослойная ткань, состоящая из пробки, феллогена и феллодермы.
Газообмен осуществляется через чечевички — разрывы в покровной ткани, заполненные рыхло расположенными клетками. На ветках деревьев и кустарников они образуют бугорки со щелью.
Корка
Корка — образуется у многих деревьев на поверхности ствола в результате многократного заложения новых прослоек перидермы во всё более глубоких слоях коры. В результате этого наружные живые ткани изолируются пробкой от центральной части стебля, деформируются и отмирают.
На этой странице материал по темам:
Ткани растений и их виды доклад краткий
Реферат на тему цитологические особенности покровных тканей растений
Покровная ткань ветки бузины
Цитологические особенности перидермы
Тест по гистологии растительные ткани
Вопросы по этому материалу:
Ботаника-наука о растенях.
Отлич.черты раст.клетки
Пластиды
3.Тканями
Образовательные ткани.
Ботаника-наука о растенях.
Ботаника охватывает широкий круг проблем: закономерности внешнего и внутреннего строения (морфология и анатомия) растений, их систематику, развитие в течение геологического времени (эволюция) и родственные связи (филогения), особенности прошлого и современного распространения по земной поверхности (география растений), взаимоотношения со средой (экология растений), сложение растительного покрова (фитоценология, или геоботаника), возможности и пути хозяйственного использования растений (ботаническое ресурсоведение, или экономическая ботаника).
По объектам исследования в ботанике выделяют фикологию (альгологию) - науку о водорослях, микология - о грибах, лихенологию - о лишайниках, бриологию - о мхах и др.; изучение микроскопических организмов, преимущественно из мира растений (бактерий, актиномицетов, некоторых грибов и водорослей), выделяют в особую науку - микробиологию. Болезнями растений, вызываемыми вирусами, бактериями и грибами, занимается фитопатология.
Ботаника пользуется как наблюдением, так и сравнительным, историческим и экспериментальным методами, включающими сбор и составление коллекций, наблюдение в природе и на опытных участках, эксперимент в природе и в условиях специализированных лабораторий, математическую обработку полученной информации. Наряду с классическими методами регистрации тех или иных признаков изучаемых растений используется весь арсенал современных химических, физических и кибернетических методов исследования.
Отлич.черты раст.клетки
Природа и основные функции веществ различны. Главнейшие из этих веществ: простые белки; некоторые углеводы, в частности глюкоза, сахароза и крахмал или близкий к нему инулин, а также целлюлоза, запасные жиры и жироподобные вещества - соединения первичного метаболизма; продукты вторичного метаболизма - танниды, полифенольные соединения, алкалоиды, изопренпроизводные и др.относится также обычный во многих растениях оксалат кальция Почти все вещества независимо от их природы в той или иной мере могут вновь вовлекаться в процессы активного метаболизма клетки.Важнейшая группа - запасные вещества. Это белки,углеводы (исключая целлюлозу) и жиры.Целлюлоза и близкие к ней по химической природе гемицеллюлозы и пектиновые вещества - структурные компоненты клеточной стенки. Вторичные метаболиты выполняют различные функции, но главнейшая из них связана с "организацией" защиты от поедания растений животными, внедрения болезнетворных микроорганизмов и т.д. Таким образом, они являются группой защитных веществ. Оксалат кальция чаще всего рассматривается в качестве конечного продукта метаболизма, формы "захоронения" ненужных или вредных клетке веществ. В технике, пищевой промышленности и медицине широко используются крахмал, глюкоза и сахароза. Танниды, или дубильные вещества, соединения полифенольные и изопренпроизводные - основа для получения большинства медицинских препаратов растительного происхождения. Пектиновые вещества широко применяются в кондитерской промышленности, а также в медицине. Некоторые из эргастических веществ крайне ядовиты. Чаще это алкалоиды, некоторые гликозиды, полипептиды (у бледной поганки). Значение запасных веществ для фармации и медицины.
Крахмал, в медицине и фармации для приготовления присыпок, паст (густых мазей), а также при производстве таблеток. Целлюлоза, велико и промышленное значение целлюлозы – из этого вещества изготовляют хлопчатобумажные ткани и бумагу.Клетки выделительных тканей по форме обычно паренхимные и тонкостенные. Они долго остаются живыми, выделяя секрет. Клетки-идиобласты по мере накопления большого количества секрета лишаются протопласта и стенки их нередко опробковевают.Ткани внутренней секреции могут быть представлены отдельными клетками-идиобластами, вместилищами выделений, смоляными ходами, эфирномасляными каналами и млечниками. Пластиды - бесцветные или окрашенные тельца в протоплазме растительных клеток, представляющие собой сложную систему внутренних мембран и выполняющие различные функции. Бесцветные пластиды называют лейкопластами, различно окрашенные - хромопластами, зеленые - хлоропластами.
Только в растительных клетках присутствуют пластиды. Эти мембранные органеллы в зависимости от окраски можно разделить на лейкопласты, хромопласты и хлоропласты. В клетке высших растений содержится около 40 хлоропластов в которых происходит фотосинтез. Как и митохондрии, хлоропласта содержат собственную ДНК и рибосомы. Они способны к автономному размножению, не зависящему от деления клетки. В зависимости от окраски, связанной с наличием или отсутствием тех или иных пигментов, различают три основных типа пластид:
Хлоропласты (зеленого цвета), - хромопласты (желтого, оранжевого или красного цвета) и - лейкопласты (бесцветные).
Самые большие из них - хлоропласты - достигают у высших растений 4-10 мкм длины и хорошо различимы в световой микроскоп. Хлоропласты встречаются во всех зеленых органах растений, лейкопласты весьма обычны в клетках органов, скрытых от солнечного света, - корнях, корневищах, клубнях, а также в ситовидных элементах некоторых покрытосеменных. Хромопласты содержатся в клетках лепестков многих растений, зрелых окрашенных плодах (томаты, шиповник, рябина), иногда - в корнеплодах (морковь).
Строение пластид может быть рассмотрено на примере хлоропластов. Они имеют оболочку, образованную двумя мембранами: наружной и внутренней. Внутренняя мембрана вдается в полость хлоропласта немногочисленными выростами. Мембранная оболочка отграничивает от гиалоплазмы клетки матрикс хлоропласта, так называемую строму. Как строма, так и выросты внутренней мембраны формируют в полости хлоропласта сложную систему мембранных поверхностей, отграничивающих особые плоские мешки, называемые тилакоидами, или ламеллами. Группы дисковидных тилакоидов связаны друг с другом таким образом, что их полости оказываются непрерывными. Эти тилакоиды образуют стопки (наподобие стопки монет), или граны. Тилакоиды стромы объединяют граны между собой. В мембранах тилакоидов сосредоточен главнейший пигмент зеленых растений - хлорофилл и вспомогательные пигменты - каротиноиды. Внутренняя структура хромопластов и лейкопластов проще. Граны в них отсутствуют.
В строме хлоропластов содержатся ферменты и рибосомы, отличающиеся от рибосом цитоплазмы меньшими размерами. Часто имеются один или несколько небольших зерен первичного ассимиляционного крахмала. Генетический аппарат хлоропластов автономен, они содержат свою собственную ДНК.
Основная функция хлоропластов - фотосинтез. Центральная роль в этом процессе принадлежит хлорофиллу, точнее - нескольким его модификациям. Световые реакции фотосинтеза осуществляются преимущественно в гранах, темновые - в строме хлоропласта. И хлоропласты, и митохондрии способны синтезировать собственные белковые молекулы, так как обладают собственной ДНК.
Помимо фотосинтеза, в хлоропластах осуществляется синтез АТФ и АДФ (фосфорилирование), синтез и гидролиз липидов, ассимиляционного крахмала и белков, откладывающихся в строме.
В лейкопластах пигменты отсутствуют, но здесь может осуществляться синтез и накопление запасных питательных веществ, в первую очередь крахмала, иногда белков, редко жиров. Очень часто в лейкопластах формируются зерна вторичного запасного крахмала.
Красноватая или оранжевая окраска хромопластов связана с присутствием в них каротиноидов. Считается, что хромопласты - конечный этап в развитии пластид, иначе говоря, это стареющие хлоропласты и лейкопласты. Наличие хромопластов отчасти определяет яркую окраску многих цветков, плодов и осенних листьев.
3.Тканями
называются группы клеток, которые имеют одинаковое происхождение, сходное строение и выполняют в организме одинаковую функцию.
Каждый тип ткани отличается характерным строением клеток, строго соответствующим той функции, которую данная ткань выполняет.
В соответствии с формой, строением и характером соединения клеток ткани могут быть рыхлые и плотные, живые и мертвые, паренхимные и прозенхимные. Все разнообразие растительных тканей принято подразделять на 6 главных групп: 1) образовательные, или меристемы, 2) основные, 3) покровные, 4) механические, 5) проводящие и 6) выделительные.Образовательные ткани.
Меристемы или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный рост растения. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей.
Первичные меристемы способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие "вновь" возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные,боковые, или латеральные, и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные - преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега.
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными - камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Покровные ткани, классификация, функции.
Они предохраняют органы растения от высыхания, от температурных воздействий, механических повреждений и других неблагоприятных воздействий окружающей среды. При изменениях возраста органов и их функций, покровные ткани закономерно сменяют одна другую. По происхождению различают первичные (эпидерма, ризодерма, веламен), вторичные (перидерма) и третичные (корка или ритидом) покровные ткани.
Первичные покровные ткани Эпидерма образуется из поверхностного слоя апикальной меристемы - протодермы. Она покрывает листья, плоды, части цветка и молодые стебли. Кроме защитной функции, эпидерма регулирует процессы транспирации и газообмена, принимает участие в синтезе различных веществ и др. В состав эпидермы входит несколько морфологически различных клеток: основные клетки эпидермы, замыкающие и побочные клетки устьиц, трихомы (выросты эпидермы). Клетки эпидермы живые, имеют ядра, лейкопласты, вакуоли, хлоропласты (только в замыкающих клетках устьиц). Эпидерма у большинства растений однослойная, реже многослойная. Клетки первичной покровной ткани плотно примыкают друг к другу, и не имеют межклетников. С наружной стороны вся эпидерма покрыта сплошным слоем кутикулы (прерывается только над устьичными щелями).
Ризодерма (эпиблема) образована апикальной меристемой корня. Она покрывает молодые корневые окончания и именно через ризодерму происходит поглощение воды и минеральных солей из почвы. Кроме того, она взаимодействует с микроорганизмами почвы, из корня в почву выделяются вещества, помогающие почвенному питанию. Клетки ризодермы имеют очень тонкие оболочки. У первичной покровной ткани корня нет кутикулы, вследствие чего эти клетки имеют оболочки легко проницаемые для воды. На небольшом расстоянии от кончика корня образуются корневые волоски - выросты ризодермы.
Веламен, как и ризодерма, происходит из поверхностного слоя апикальной меристемы корня. Эта своеобразная ткань покрывает корни эпифитов и некоторых других растений, приспособленных к жизни на периодически пересыхающих почвах (аспидистра, аспарагус, алоэ, кливия). Веламен от ризодермы отличается многослойностью. Протопласт веламена отмирает и поэтому всасывает воду не осмотическим, а капиллярным путем.
Эмергенцы (нем., единственное число Emergenz, от лат. emerge - появляюсь, выхожу), выросты на поверхности стеблей и листьев растений, образованные не только эпидермисом, но и глубже лежащими тканями (в отличие от волосков ). Это - жгучие волоски крапивы, шипы на стеблях розы и на плодах дурмана, железки на листьях росянки и др.Вторичная покровная ткань - Перидерма. Перидерма сложная, многослойная вторичная покровная ткань стеблей и корней многолетних(реже однолетних) растений 1 . Перидерма сменяет первичные покровные ткани осевых органов, которые постепенно отмирают и слущиваются. У большинства двудольных и голосемянных перидерма обычно появляется в течение первого вегетативного периода в тех частях корня и побега, которые прекратили рост в длину. В основе перидермы стебля лежит вторичная меристема - феллоген. Феллоген стебля чаще возникает из клеток основной паренхимы, лежащей под эпидермой и сохранившей слабую меристематическую активность. У корней основой образования феллогена может быть перицикл. Феллоген может закладываться отдельными участками, позднее сливающимися, либо сразу по всей окружности осевого органа. Прежде всего образуется инициальный ряд уплощенных по форме таблитчатых клеток. Позднее инициальные клетки периклинально делятся, формируя два ряда меристематических клеток. Внутренний ряд дифференцируется как феллодерма (живые паренхим-ные клетки) и далее не делится. Наружный - в ходе серии периклинальных делений формирует различное количество слоев клеток пробки, длительное время, сохраняя меристематическую активность. Феллоген способен делиться также и антиклинально, обеспечивая правильное соотношение тканей в разрастающихся органах. Пробка, феллоген и феллодерма образуют перидерму. По мере того как формируется перидерма, зеленый цвет побегов переходит в бурый. «Вызревшие» к осени побеги первого года способны выдержать зимние морозы. Пробка состоит из отмерших клеток, лишенных межклетников. Их оболочки пропитаны суберином. Клетки пробки воздухо- и водонепроницаемы. Многослойная пробка образует защитный футляр, предохраняющий живые ткани от потери влаги, от резких температурных колебаний и проникновения болезнетворных микроорганизмов. На стволах и сучьях некоторых деревьев (пробковый дуб, бархат амурский) формируется мощный слой пробки, достигающий нескольких сантиметров толщины. Такую пробку используют для промышленного получения укупорочных материалов. Живые ткани, лежащие под пробкой, нуждаются в газообмене и удалении избытка влаги. Поэтому в перидерме довольно рано образуются чечевички - отверстия, «заполненные» рыхлой выполняющей тканью, состоящей из закругленных паренхимных слабоопробковевших клеток с многочисленными межклетниками. Через чечевички осуществляется «проветривание» стебля. Чечевички, Имеющие вид небольших бугорков различной формы, хорошо заметны на поверхности молодых побегов деревьев и кустарников. Чечевички в виде мелких крапин заметны и на некоторых плодах, например яблоках и грушах. По мере утолщения побегов форма чечевичек меняется. На стволах березы их остатки заметны в виде характерных поперечных полосок и черточек. Перидерма образуется, как сказано, у двудольных и хвойных растений. У однодольных (пальмы, драцены, алоэ) иногда путем многократного деления и опробковения периферических клеток первичной коры образуется защитная ткань функционально сходная с перидермой. В многолетних осевых органах двудольных и хвойных растений обычно развивается несколько перидерм. Каждая последующая закладывается глубже предыдущей. Со временем живые клетки наружных перидерм и располагающихся между ними тканей отмирают, образуя мощный покровный комплекс - корку.Она формируется на стволах, многолетних ветвях и корнях деревьев. В отличие от более или менее гладкой перидермы корка нередко трещиноватая и неровная (чешуйчатая пробка). Существует, правда, и относительно гладкая, так называемая кольцевая корка (например, у винограда). Эти различия связаны с особенностями заложения внутренних перидерм. Толстая корка надежно предохраняет стволы деревьев от механических повреждений, лесных низовых пожаров, резкой смены температур, ожогов прямыми солнечными лучами, проникновения патогенных микроорганизмов. Обычно у древесных пород корка начинает закладываться в зрелом возрасте: у сосны в 8-10 лет, у дуба в 25-30 лет. Ежегодно наращиваясь, с годами она может достигать значительной толщины.
Первичная покровная ткань.
Кожицу листьев и стеблей называют эпидермой
, кожицу корня – эпиблемой
.
Основные функции эпидермы – защита молодых органов от высыхания, механическая защита и газообмен. Эпидерма
, как правило, представлена одним слоем плотно сомкнутых
клеток, на внешней поверхности жироподобное вещество кутин образует защитную пленку – кутикулу. На поверхности кутикулы часто имеется восковой налет. Стенки клеток обычно извилистые, наружные стенки толще остальных.
Зпиблема, ризодерма (от греческого epiblema - покрывало, derma - кожа и rhiza - корень) - первичная покровная ткань корня, выполняющая функции защиты и поглощения питательных веществ. На поверхности клеток образуются боковые выросты – корневые волоски. Устьица и кутикула у эпиблемы отсутствуют.
Экзодерма (от экзо... и греч. derma - кожа) - часть первичной коры, корня, одно- или многоклеточное образование, находящееся непосредственно под первичной покровной тканью - эпиблемой . Особенно характерна для однодольных растений, у которых со временем заменяет эпиблему. Клетки экзодермы остаются живыми, хотя их стенки опробковевают. В экзодерме иногда встречаются пропускные клетки, хотя есть данные, что транспорт веществ может осуществляться и непосредственно через экзодерму.
Для газообмена и транспирации в эпидерме имеются специальные образования – устьица
.
Устьице
представляет собой щелевидное отверстие в эпидерме, ограниченное двумя клетками бобовидной формы. Это замыкающие клетки. В отличие от остальных клеток эпидермы они содержат хлоропласты
. Стенки замыкающих клеток, обращенные в сторону устьичной щели, утолщены. Клетки эпидермы, окружающие замыкающие, называют побочными или прилегающими. Под устьицем находится газовоздушная камера. Замыкающие и побочные клетки, устьичная щель и газовоздушная камера образуют устьичный аппарат. Устьица чаще
располагаются на нижней стороне листа
.
Иногда клетки эпидермы образуют различные придатки, волоски и чешуйки (трихомы). Волоски выполняют защитную функцию, сильное опушение защищает растение от перегрева и потери влаги. Железистые волоски выполняют защитную функцию (например, у крапивы).
Вторичная покровная ткань
, перидерма
.
Перидерма
(от пери... и греч derma - кожа) - вторичная покровная ткань стеблей и корней, корневищ и клубней, редко листьев и плодов. Иногда рассматривается как комплекс тканей. Образуется из вторичной меристемы - феллогена. Состоит из:
феллемы
– собственно пробки
,
феллогена
– пробкового камбия
и
феллодермы
– пробковой паренхимы (внутренняя живая ткань)
…
Из-за изолирующего многослойного пробкового покрова перидерма непроницаема для воды и воздуха. Обмен с внешней средой осуществляется через чечевички - прорывы в перидерме, образующиеся над устьицами. Перидерма сменяет эпидерму, которая постепенно отмирает и слущивается. Закладывается преимущественно в стеблях и корнях.
Третичная покровная ткань
Ткань, образующаяся из вторичной ткани в результате происходящих в ней новообразовательных процессов (напр., заложение прослоек камбия в глубже расположенной вторичной ткани).
К третичной ткани относится ребристая корка
на поверхности толстых стволов древесных растений.
Корка (ритидом)
, третичная покровная ткань. При образовании корки новый слой феллогена и перидермы закладывается в основной ткани, лежащей глубже первой наружной перидермы. Вновь образовавшиеся слои пробки отчленяют к периферии органа не только перидерму, но и часть лежащей под ней паренхимы коры. Так возникает толстое многоклеточное и мертвое образование. Так как корка не может растягиваться, при утолщении ствола она лопается и образуются трещины.
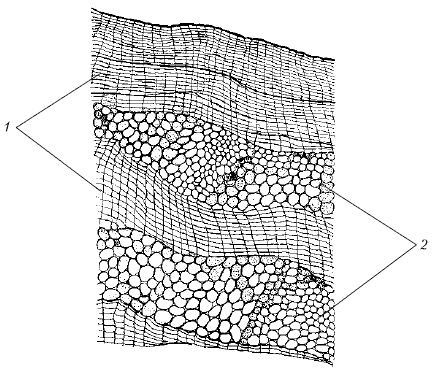
Корка (ритидом)
образуется у многолетних растений в корне, стебле, корневище.
Каждый год в более глубоких слоях закладывается новый слой феллогена и образуется перидерма. Наружный слой перидермы - пробка - изолирует все вышележащие ткани, в результате чего они отмирают. Таким образом, совокупность многочисленных перидерм с отмершими между ними тканями и является коркой.
Под покровной тканью следует понимать комплекс клеток, расположенных снаружи всех органов растения.
При попытке дать покровной ткани более точное определение, соответствующее характеру каждого отдельного случая, (встречается ряд трудностей. Покровные ткани различных органов весьма разнообразны по строению и по функциям. Покровные ткани надземных органов - листа, стебля - служат для защиты от высыхания и повреждения внутренних, более нежных тканей, выполняют выделительную функцию, при их посредничестве осуществляется газообмен с окружающей средой и пр. Покровы же корней, особенно окончаний их, поглощают воду с растворенными в ней минеральными веществами, содействуют растворению некоторых твердых частиц почвы и защищают внутренние ткани корня от механических повреждений. Покровы осевых органов у многолетних растений к концу первого вегетационного периода в большинстве случаев начинают сильно изменяться: покровы первого года вегетации отмирают, а из них или из других тканей, расположенных в непосредственной близости, образуется покровная ткань другого типа как по структуре, так и по ряду дополнительных функций. Следовательно, необходимо различать первичные покровные ткани и вторичные покровные ткани . Последние или возникают вновь, или представляют собой видоизменение структуры первичной ткани.
Первичная покровная ткань всех растительных органов называется кожицей, или эпидермисом .
Эпидермис образуется из поверхностного слоя первичной меристемы, называемого протодермой . Эпидермис состоит из клеток, как правило, изодиаметрического характера и всегда с живым содержимым. При отмирании протопласта эпидермальных клеток последние перестают служить покровом и защитой находящимся за ними тканям и заменяются другой покровной тканью.
Если рассматривать эпидермальные клетки снаружи, то можно убедиться, что очертания их в разных органах и у разных растений весьма различны. Очертания эпидермальных клеток формируются в большой зависимости от соотношения в скорости роста органа в целом и его поверхности. Например, клетки эпидермиса листа с широкой пластинкой, одинаково разрастающейся как в длину, так и в ширину, имеют извилистые очертания; а клетки эпидермиса стебля молодого растения, сильно вытягивающегося в длину, значительно более вытянуты и имеют менее извилистые стенки, чем клетки эпидермиса листа того же растения. Линейные листья однодольных растений отличаются длинными эпидермальными клетками.
Клетки эпидермиса весьма прочно соединены друг с другом, значительно прочнее, чем с клетками подстилающей эпидермис ткани. Вследствие этого эпидермис часто легко отрывается от прочих тканей, но сам не разрывается и сдирается относительно большими лоскутами. Эпидермис обычно не имеет межклетников, но бывают и исключения. Например, межклетники встречаются в эпидермисе лепестков. Кроме того, в эпидермисе других надземных, преимущественно зеленых, органов имеются щелевидные межклетники, окруженные специальными замыкающими их клетками, - устьица .
В молодых эпидермальных клетках почти вся полость клетки заполнена протопластом с отчетливо видным округлым клеточным ядром и достаточно заметными, хотя и мелкими, лейкопластами. По мере разрастания клетки и увеличения размеров вакуолей цитоплазма занимает постенное положение, и ядро, если оно расположено не в центре клетки, а в цитоплазме, прижатой к оболочке, сплющивается и вытягивается.
Во вполне выросшей эпидермальной клетке вакуоля занимает почти всю клетку, она наполнена клеточным соком, бесцветным или окрашенным растворенными в нем пигментами. Протопласт в виде очень тонкого слоя, нередко с трудом различимого, прижат к оболочке. Часто в такой клетке пластиды разрушаются, совершенно исчезая. Это наблюдается преимущественно у светолюбивых травянистых растений. Впрочем и у таких растений иногда в эпидермальных клетках нижней стороны листа есть хлорофилловые зерна. У растений, произрастающих в тени, хлоропласты часто имеются во всех эпидермальных клетках. У некоторых растений пластиды в эпидермисе листьев сохраняются в виде мелких шаровидных лейкопластов, группирующихся вокруг клеточного ядра.
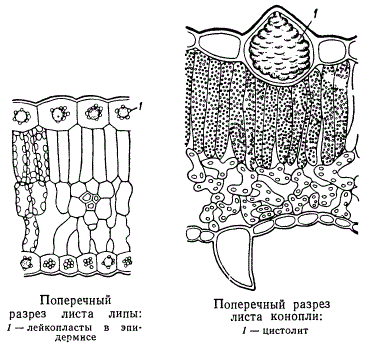
В эпидермальных клетках образуются различные продукты жизнедеятельности протопласта. Особенно своеобразны цистолиты. , они представляют собой весьма объемистые отложения углекислой извести, возникающие в гипертрофически разросшихся клетках эпидермиса (идиобластах, в данном случае - литоцистах). Если подействовать на эти цистолиты кислотой, например, соляной, то основная масса цистолита растворится и останется лишь целлюлозная основа, прикрепленная к оболочке клетки, на которой откладывается углекислый кальций. Форма и размеры цистолитов у различных растений различны. Они свойственны определенным видам растений. В нашем примере приводится цистолит в листьях конопли. Классическим примером служат цистолиты в листьях фикуса.
Оболочка клеток эпидермиса, особенно наружная стенка их, часто пропитывается или солями кальция, или соединениями кремния (злаки, хвощи). На поверхности эпидермальных клеток очень часто выделяются кутикула и кутикулярные слои. Кутикула затягивает всю поверхность сформировавшегося эпидермиса, прерываясь только устьичными щелями. Кроме кутикулы, снаружи ее у большинства растений выделяется воск в виде тонкого мелкозернистого налета. У некоторых тропических растений восковой покров на листьях достигает значительной толщины (0,5 см). И кутикула и восковой налет не смачиваются водой и предохраняют ткани, расположенные под ними, от потери воды.
Условия произрастания определяют некоторые особенности структуры эпидермальных тканей растения. Эпидермис листьев и стеблей водных растений почти не имеет кутикулы и тем более воскового налета, а у растений засушливых местообитаний кутикула, как правило, образуется. Оболочки эпидермальных клеток надземных органов некоторых растений одревесневают, становятся весьма толстыми, с сильно сократившимися размерами клеточных полостей. При этом часто одревесневают не только клетки самого эпидермиса, но и подстилающие его клетки. Поверхность эпидермиса надземных органов очень редко остается гладкой и однородной в течение всего времени существования этой ткани. Даже на кутикуле почти всегда возникают бугорки, складки, штрихи, придающие поверхности различный, иногда довольно прихотливый рисунок. Кроме того, клетки эпидермиса часто образуют разнообразнейшие выросты - волоски . Волоски бывают одноклеточными и многоклеточными, ветвистыми и неветвистыми. Форма волосков также варьирует: существуют волоски звездчатые, головчатые, в виде чешуек, прицепок и др. Форма, размеры и распределение волосков на поверхности органов характерны для определенных групп растений, и эти признаки часто служат систематическими.
Обычно волоски отделяются перегородкой от той эпидермальной клетки, на которой они образовались. Но есть волоски, представляющие собой даже во вполне развитом состоянии не более как выросты клеток эпидермиса, не отделяющиеся от них специальной перегородкой. Если такие выросты эпидермальных клеток очень короткие, то их называют сосочками . Сосочки особенно распространены на лепестках цветков, придавая им бархатистость.
Многоклеточные волоски нередко чрезвычайно сильно разрастаются, составляющие же их клетки одревесневают и волоски становятся твердыми. Такие волоски называют шипами (малина, старые стебли тыквы, стебли некоторых сортов розы, барбариса и др.).
Все многообразие волосков, руководствуясь физиологическим принципом, можно свести к двум основным типам: кроющие и железистые . Очень часто волоски обоих типов существуют на одном и том же органе. Например, на рисунке изображены простые кроющие и железистые волоски прицветной чешуйки рудбекии.
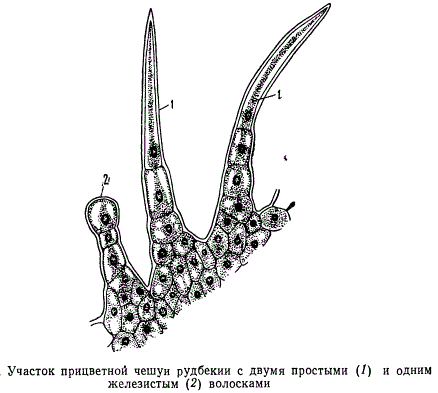
Кроющие волоски остаются живыми сравнительно недолго. Когда протопласт волоска отмирает, остов его, состоящий из оболочки, сохраняется, наполняясь воздухом. Обычно эти наполненные воздухом блестящие белые остовы волосков и образуют столь характерный волосяной покров тела растений.
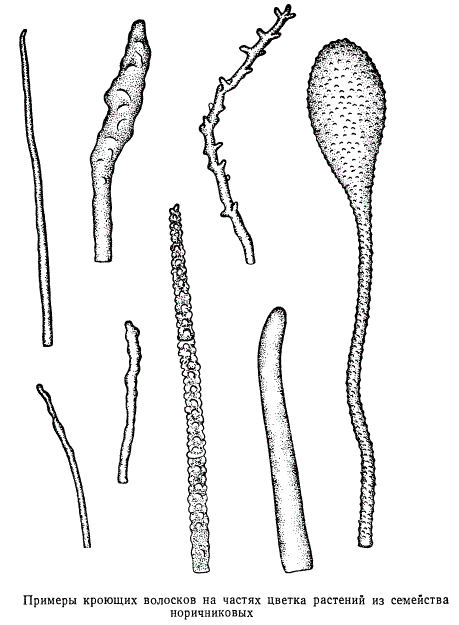
Морфологическое разнообразие кроющих волосков представлено на рисунке, на котором изображены волоски частей цветка растений из семейства норичниковых. Все волоски за исключением двух одноклеточные.
Железистые волоски сохраняют живое содержимое своих клеток несколько дольше кроющих. Протопласт клеток железистых волосков, особенно верхушечных, образующих головку волоска клеток, нередко выделяется значительной вакуолизацией и почти всегда содержит крупное клеточное ядро. Если протопласт клеток железистого волоска разрушается, то нежные оболочки их спадаются и волосок, засохнув, отпадает, не оставляя после себя почти никакого следа. Морфология железистых волосков так же, как и кроющих, беспредельно разнообразна. Обычно выделения железистых волосков состоят или из смолистых веществ, или же из эфирных масел, сахаристых веществ, слизей, а иногда просто из воды и растворенных в ней веществ различной природы.
Железистые волоски, покрывающие, например, почечные чешуйки и кроющие листья почек, выделяют смолистое вещество, как бы закупоривающее почку и предохраняющее нежные листочки от высыхания. Возможно, что выделения железистых волосков являются конечными продуктами сложных биохимических процессов, протекающих в растительных организмах.
Железистые волоски, выделяющие воду и имеющие специально приспособленную для этого структуру, называют гидатодами .
Среди железистых волосков особенно своеобразны по своей организации так называемые жгучие волоски, общеизвестным примером которых являются жгучие волоски крапивы. Стенки волоска крапивы пропитаны соединениями кремния, вследствие чего становятся очень ломкими, особенно у верхушки. Такой волосок обламывается от легчайшего соприкосновения, причем кончик волоска превращается в острую иглу, легко вонзающуюся в кожу животного или человека. Такие же волоски с еще более жгучим секретом образуются на различных частях Alonsoa. В большинстве других случаев оболочки железистых волосков нежные, даже если они и выделяют раздражающий кожу секрет, как это имеет место, например, у некоторых видов борщевика.
Значение волосков разнообразно и во многих случаях еще не выяснено. Принято считать, что наиболее распространенной функцией волосков типа кроющих является предохранение поверхности тела растения от излишней потери воды. Блестящие белые волоски, с одной стороны, отражают прямые солнечные лучи, с другой - густым сплетением создают над эпидермисом среду, способствующую уменьшению транспирации. Однако специальными опытами по крайней мере в отношении частей цветка показано, что покрывающие их жизнедеятельные волоски увеличивают интенсивность транспирации.
Обычно ознакомление с различными волосками основано на изучении волосков, покрывающих уже выросшие органы. За исключением железистых волосков, описываются волоски мертвые, лишенные живого содержимого. В живом состоянии такие волоски, несомненно, более связаны с процессами жизнедеятельности растительного организма и играют иную физиологическую роль, чем отмершие сухие волоски. Подтверждением этому положению является развитие волосков на колосках злаков. На молодых цветочных и колосковых чешуях колоса, когда он еще находится в стадии так называемой трубки и окончательно еще не оформился, по краям чешуй, вдоль средней жилки и на молодой ости, если колос будет остистым, расположены волоски с хорошо выраженным крупным клеточным ядром и густой цитоплазмой. Эти волоски пребывают в наиболее активном жизненном состоянии. Из всех частей колоска колосковая и цветковая чешуи развиваются в первую очередь, а волоски на них формируются раньше всех прочих клеток, составляющих ткани этих чешуй. Волоски на чешуях колоска остаются в деятельном состоянии во все время формирования завязи и зародышевого мешка в ней. Как только зародышевый мешок готов к оплодотворению или, в других случаях, когда оплодотворение произойдет, содержимое волосков начинает быстро отмирать. Волоски, расположенные вдоль средней жилки и ости, успевшие значительно разрастись, превращаются в шипики и бугорки, претерпевают одревеснение и придают колосу хорошо известную на ощупь шероховатость.
Таким образом, волоски на начинающем развиваться колоске злака в своем деятельном состоянии связаны с важнейшими процессами в жизни растения - формированием и созреванием полового аппарата, во время которых идет активнейший обмен веществ.
Форма волосков на молодых колосковых чешуях разнообразна.

На молодых завязях различных растений часто образуется много волосков, в содержимом которых имеется алкалоид или какое-либо другое физиологически активное вещество. Все эти волоски в период формирования завязи находятся в жизнедеятельном состоянии, а после оплодотворения начинают отмирать и отпадают. На рисунке изображены железистые волоски завязи девясила; позднее, после оплодотворения, они быстро исчезают. Ясно, что волоски, столь хорошо и в изобилии развивающиеся на поверхности завязи и отпадающие после оплодотворения, в своем развитии связаны с интенсивно идущими в завязи процессами формирования женских половых элементов.

На семянках одной из ромашек, произрастающих в пустыне, можно найти своеобразные волоски, изображенные на рисунке. Каждый волосок состоит из двух морфологически хорошо отличающихся частей: основания, состоящего из нескольких члеников - отдельных клеток, и верхушки, занятой одной ослизняющейся клеткой. Каждый членик основания снабжен целлюлозными тяжами утолщений оболочки, образующими изящный переплет - подобие каркаса. На рисунке все три волоска представляют собой различные стадии ослизнения конечной клетки. Ослизняющиеся клетки и волоски на поверхности плодов некоторых растений, произрастающих в пустынных местностях, обособляются относительно очень рано, они хорошо различимы на далеко еще не зрелых плодах. Часто такие клетки в виде коротких толстых волоскоподобных образований располагаются на верхушках ребер семянок, причем при посредстве особых клеток, называемых гидроцитами , оболочки которых снабжены разнообразными утолщениями, наружные волоскоподобные клетки сообщаются с сосудистыми пучками.
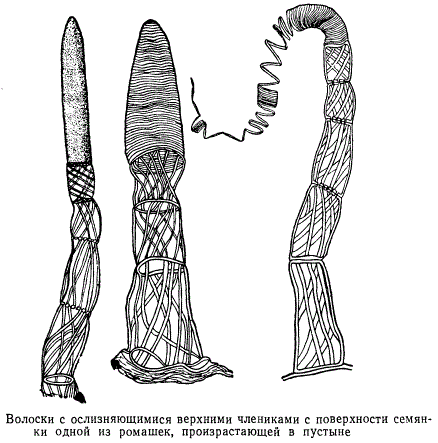
Таким образом, создается своеобразная гидроцитная система, играющая большую роль в водном режиме созревающего плода.
По созревании семянки волоскоподобные гигроскопические образования засыхают и остаются на поверхности в виде небольших бугорков и бородавочек или опадают, или же превращаются в ослизняющиеся клетки.

Эпидермис образуется не только снаружи органа, но и внутри, если в органе формируется специальная полость. Например, ткань, выстилающая полость плодов, по своему происхождению идентична эпидермису, покрывающему плоды снаружи. В связи с нахождением покровной ткани внутри полости, где влажность воздуха и прочие условия отличаются от условий внешней среды, особенности структуры внутреннего эпидермиса плода несколько иные, чем наружного эпидермиса.
Клетки внутреннего эпидермиса плода гороха соединены между собой рыхло, кутикула или очень тонка или отсутствует. Значительная часть клеток вытянута в волоски различной длины с тонкими оболочками. Местами и особенно около семян образуется густой пушок из сосочкоподобных волосков, облекающий семя со всех сторон. То же наблюдается в плодах конских бобов, только покров, снабженный волосками, отличается более густым и равномерным скоплением последних.
Несомненно, образование таких сосочкоподобных волосков, соприкасающихся с развивающимися семенами, имеет какое-то физиологическое значение в процессе развития семян.
У некоторых растений оболочки внутреннего эпидермиса плода довольно значительно утолщаются и иногда одревесневают, особенно ко времени окончательного созревания плода. Хорошим примером такого строения внутреннего эпидермиса плода является эпидермис, покрывающий изнутри стенки коробочек мака. Волоски в таких случаях развиваются не всегда и никогда не достигают значительной длины; кутикула выражена более заметно, поверхность эпидермиса часто бывает гладкой, клетки имеют утолщенные оболочки и плотно сомкнуты между собой, иногда среди них встречаются устьица.
Таким образом, роль волосков и подобных им образований в жизни растительных организмов весьма разнообразна и меняется в зависимости от условий произрастания и возраста растений.
Продолжительность существования эпидермиса на надземных побегах невелика и обычно ограничивается одним вегетационным периодом. Но есть исключения и не столь редкие, особенно у растений более южных районов. Например, листья лавровишни и лавра существуют больше одного года, следовательно, и эпидермис их сохраняется больше одного года. Есть деревья, у которых эпидермис на побегах сохраняется несколько лет, даже на толстых стволах многолетних деревьев. Примерами служат эвкалипт, платан, стеркулия, а на севере - серая ольха, черемуха и др. Многолетен эпидермис также на хвое сосны, ели, туи и др.
На поверхности многолетних эпидермисов развивается относительно толстая кутикула. У хвойных оболочки эпидермальных клеток одревесневают.
В отличие от надземных частей растений ткань, покрывающая молодые части корней, лишена кутикулы вследствие чего клетки этой ткани имеют оболочки, легко проницаемые для воды. Кроме того, у большинства растений при соответствующих условиях клетки, покрывающие конечные участки корней, образуют тонкие выросты, не отделяющиеся от образовавших их клеток перегородками. Эти клетки называются корневыми волосками . Корневые волоски у различных растений бывают различной длины, но в общем длина их колеблется от 0,15 до 8,0 мм. На корнях проростков гороха, выращенных во влажной атмосфере, длина корневых волосков равна 2,5 мм, у конских бобов - 0,8 мм. У одного и того же растения в зависимости от условий среды, в которой развиваются корни, длина корневых волосков может значительно колебаться. В более сухих условиях корневых волосков образуется больше, чем во влажных; у растений, произрастающих в почве, покрытой водой, корневые волоски часто совершенно не образуются.
Оболочка корневых волосков целлюлозная, очень нежная, кутикула на жизнедеятельных волосках не образуется. Наиболее обычна цилиндрическая форма корневых волосков. Ветвистые корневые волоски формируются лишь у редких растений, но при слипании с частицами почвы строгость цилиндрической формы их нарушается. По существу, корневые волоски представляют собой выросты клеток эпидермиса, нередко сильно удлиненные. Однако у немногих растений корневые волоски все же отделяются перегородкой от породившей их эпидермальной клетки.
Начинают развиваться корневые волоски не на самом кончике корня, а на некотором расстоянии от него, примерно на 0,5-3,0 мм от верхушки. Зона, занимаемая корневыми волосками, по длине корня не очень велика, причем длина зоны зависит от многих условий, но не превышает нескольких миллиметров. Корневые волоски недолговечны: достигнув некоторого предела существования, они отмирают, сморщиваются и отпадают. Отмирание корневых волосков происходит в том конце занимаемой ими зоны, где расположены наиболее старые волоски, а на противоположном полюсе зоны происходит новообразование волосков. Корневые волоски постоянно возникают вновь в более молодых участках кончика корня и отмирают в более старых, структура которых становится более постоянной. Таким образом, разветвлениями корня охватываются все новые и новые частицы почвы, с которыми слипаются корневые волоски. Корневые волоски расположены на кончиках корней в занимаемой ими зоне очень густо. Например, у кукурузы на 1 мм 2 поверхности кончика корешка приходится около 425 корневых волосков, а у гороха при тех же самых условиях - 232. За счет образования корневых волосков поверхность корня сильно увеличивается. Так, поверхность покрытой корневыми волосками части кончика корня кукурузы в 5,5 раз больше поверхности такой же части корня без волосков, а у гороха - даже в 12,2 раза. Увеличение поверхности корня содействует выполнению одной из основных его функций - поглощению воды и растворенных в ней веществ.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .