Клеточная оболочка - типичный компонент растительной клетки, является продуктом жизнедеятельности протопласта.
Функции:
1. Прочные и жесткие клеточные оболочки, служат механической опорой для органов растения.
2. Оболочка ограничивает растяжение протопласта вакуолью, а размер и форма зрелой клетки перестают изменяться.
3. В наружных тканях клеточные оболочки, защищают лежащие глубже клетки от высыхания.
4. По клеточным стенкам, примыкающим к друг другу, могут передвигаться различные вещества и вода от клетки к клетке (путь через апопласт).
5. Они оказывают влияние на поглощение, транспирацию и секрецию.
Клеточные стенки, как правило, бесцветны и легко пропускают солнечный свет. Стенки соседних клеток скреплены пектиновой срединной пластинкой . Срединная пластинка - единый слой, общий для двух соседних клеток. Она представляет собой несколько видоизмененную клеточную пластинку, возникшую в процессе цитокинеза. Срединная пластинка менее обводнена, в ней могут присутствовать молекулы лигнина. Углы клеточных стенок в результате внутриклеточного давления могут округляться, и между соседними клетками образуются межклетники. Все стенки клеток растения, связанные одна с другой и примыкающие к заполненным водой межклетникам, обеспечивают существование сплошной обводненной среды, в которой свободно передвигаются водорастворимые вещества.
Строение и химический состав.
Первичная клеточная стенка.
Первоначально кнаружи от плазмалеммы возникает первичная клеточная стенка.
Состав: целлюлоза, гемицеллюлоза, пектин и вода.
Первичные клеточные стенки соседних клеток соединены протопектиновой срединной пластинкой. В клеточной стенке линейные очень длинные (несколько микрон) молекулы целлюлозы, состоящие из глюкозы, собраны в пучки - мицеллы, которые, в свою очередь, объединяются в микрофибриллы – тончайшие (1,5…4 нм) волоконца неопределенной длины, а затем в макрофибриллы. Целлюлоза образует многомерный каркас, который погружен в аморфный сильно обводненный матрикс из нецеллюлозных углеводов: пектинов, гемицеллюлоз и др. Именно целлюлоза обеспечивает прочность клеточной стенки. Микрофибриллы эластичны и по прочности на разрыв сходны со сталью. Полисахариды матрикса определяют такие свойства стенки, как высокая проницаемость для воды, растворенных мелких молекул и ионов, сильная набухаемость. Благодаря матриксу по стенкам, примыкающим к друг другу, могут передвигаться вода и вещества от клетки к клетке (путь через апопласт по «свободному пространству»). Некоторые гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ.
Рост стенки.
При делении клеток создается заново лишь клеточная пластинка. На нее обе дочерние клетки откладывают собственные стенки, состоящие главным образом из гемицеллюлозы. При этом образование стенки происходит и на внутренней поверхности остальных стенок, принадлежащих материнской клетке. Клеточная пластинка преобразуется в срединную, она обычно тонка и почти неразличима. После деления клетка вступает в фазу растяжения за счет поглощения клеткой воды и роста центральной вакуоли. Тургорное давление растягивает стенку, в которую внедряются мицеллы целлюлозы и вещества матрикса. Такой способ роста носит название интуссусцепции , внедрения. Оболочки делящихся и растущих клеток называют первичными. Они содержат воды до 90 %, в сухом веществе преобладают полисахариды матрикса: у двудольных пектины и гемицеллюлозы в равном соотношении, у однодольных – в основном гемицеллюлозы; содержание целлюлозы не превышает 30 %. Толщина первичной стенки не более 0,1…0,5 мкм.
К моменту, когда рост клетки заканчивается, рост клеточной стенки может продолжаться, но уже в толщину. Этот процесс носит название вторичного утолщения. При этом на внутренней поверхности первичной клеточной стенки откладывается вторичная клеточная стенка. Рост вторичной клеточной стенки происходит в результате аппозиции , наложения новых мицелл целлюлозы на внутреннюю поверхность клеточной стенки. Таким образом, наиболее молодые слои клеточной стенки ближе всего к плазмаллеме.
Для некоторых типов клеток (многие волокна, трахеиды, членики сосудов) образование вторичной стенки – основная функция протопласта, после завершения вторичного утолщения он отмирает. Однако это не обязательно. Вторичная стенка выполняет главным образом механические, опорные функции. В ее составе значительно меньше воды и преобладают микрофибриллы целлюлозы (40…50 % сухого вещества). Во вторичных стенках волокон льна и волосков хлопчатника содержание целлюлозы может достигать 95 %.
Механизм построения клеточной стенки. Клеточная стенка образуется в результате деятельности протопласта. В соответствии с этим вещества поступают в стенку изнутри, со стороны протопласта. Строительные материалы – молекулы целлюлозы пектина, лигнина и других веществ - накапливаются и частично синтезируются в цистернах аппарата Гольджи. Упакованные в пузырьки аппарата Гольджи, они транспортируются к плазмалемме. Разорвав ее, пузырек лопается, и содержимое его оказывается снаружи плазмалеммы. Мембрана пузырька восстанавливает целостность плазмалеммы. Благодаря ферментной активности плазмалеммы идет сборка фибрилл целлюлозы строение клеточной стенки. Образуемые плазмалеммой фибриллы накладываются изнутри, не переплетаясь. В их ориентации большая роль принадлежит микротрубочкам, располагающимся под плазмалеммой параллельно формирующимся фибриллам.
2. Поры. Видоизменения клеточной стенки.
Поры. При образовании первичной клеточной стенки в ней выделяются более тонкие участки, где фибриллы целлюлозы лежат более рыхло. Канальцы эндоплазматической цепи проходят здесь через клеточные стенки, соединяя соседние клетки. Эти участки называются первичными поровыми полями , а канальцы эндоплазматической сети, проходящие в них, - плазмодесмами .
Рост в толщину происходит у клеточной стенки неравномерно, неутолщенными остаются небольшие участки первичной клеточной стенки в местах расположения первичных поровых полей (поровых каналов). Поровые каналы двух соседних клеток располагаются обычно друг против друга и разделяются замыкающей пленкой поры - двумя первичными клеточными стенками с межклеточным веществом между ними. В пленке сохраняются субмикроскопические отверстия, через которые проходят плазмодесмы. Таким образом, пора - это два поровых канала и замыкающая пленка между ними .
Плазмодесмы пронизывают замыкающие пленки пор. В каждой клетке имеется от нескольких сотен до десятков тысяч плазмодесм. Плазмодесмы встречаются только - в растительных клетках, там, где имеются твердые клеточные стенки. Плазмодесмы образуются из канальцев ЭР, которые остаются в клеточной пластинке между двумя дочерними клетками. При воссоздании ЭР обеих клеток они оказываются соединенными через плазмодесмы.
Плазмодесма проходит через плазмодесменный канал в замыкающей пленке поры. Плазмалемма, выстилающая канал, и гиалоплазма между ней и плазмодесмой непрерывны с плазмалеммами и гиалоплазмами смежных клеток. Таким образом, протопласты соседних клеток связаны между собой каналами плазмодесм и плазмодесмами. По ним происходит межклеточный транспорт ионов и молекул, а также гормонов. Объединенные плазмодесмами протопласты клеток в растении образуют единое целое - симпласт. Транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
В процессе жизнедеятельности клетки целлюлозная клеточная стенка может претерпевать видоизменения.
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной , или межклеточной пластинкой (межклеточным веществом) (рис. 2.16 ). Срединная пластинка обычно очень тонка и почти неразличима.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. После деления клетка вступает в фазу роста растяжением, который обусловлен интенсивным осмотическим поглощением клеткой воды, связанным с образованием и ростом центральной вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Отложение новых порций материала происходит равномерно по всей поверхности протопласта, поэтому толщина клеточной стенки не уменьшается.
Стенки делящихся и растущих клеток называют первичными . Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).
Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни ( рис. 2.16).
Рис. 2.16. Паренхимная клетка с первичной стенкой.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной (рис. 2.17 ).
Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация . Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники (рис. 2.16 ). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки – ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки – поры . Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов – простые и окаймленные (рис. 2.17 ).
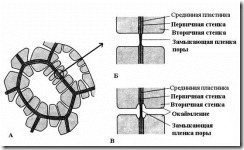
Рис. 2.17. Типы пор : А – клетки с вторичными стенками и многочисленными простыми порами; Б – пара простых пор; В – пара окаймленных пор.
У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон.
Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пор и функционирует как одно целое. Разделяющий их канал участок стенки называется замыкающей пленкой поры , или поровой мембраной . В живых клетках замыкающая пленка поры пронизана многочисленными плазмодесмами (рис. 2.18 ).
Плазмодесмы присущи только растительным клеткам. Они представляют собой тяжи цитоплазмы, пересекающие стенку смежных клеток. Число плазмодесм в одной клетке очень велико – от нескольких сотен до десятков тысяч, обычно плазмодесмы собраны в группы. Диаметр плазмодесменного канала составляет 30-60 нм. Его стенки выстланы плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы проходит мембранный цилиндр – центральный стержень плазмодесмы , непрерывный с мембранами элементов эндоплазматической сети обеих клеток. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма, непрерывная с гиалоплазмой смежных клеток.
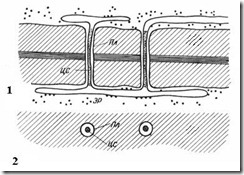
Рис. 2.18. Плазмодесмы под электронным микроскопом (схема ): 1 – на продольном срезе; 2 – на поперечном срезе; Пл – плазмалемма; ЦС – центральный стержень плазмодесмы; ЭР – элемент эндоплазматического ретикулума.
Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом , а транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
У окаймленных пор (рис. 2.17 )канал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры. Название окаймленной поры происходит оттого, что при рассмотрении с поверхности внутреннее отверстие имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде круга большего диаметра или более широкой щели.
Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, в то же время не снижая прочности клеточной стенки.
Клетки растений окружены плотной полисахаридной оболочкой, выстланной изнутри плазмалеммой.
Образование клеточной стенки происходит в метафазе и телофазе клеточного деления. В экваториальной зоне деления возникает срединная пластинка, состоящая из пектата кальция, которая, нарастая от центра к периферии, отделяет одну новообразованную клетку от другой. Срединная пластинка с той и другой стороны покрывается первичной клеточной стенкой. Рост в толщину происходит за счет наложения новых слоев со стороны содержимого каждой клетки. Рост клетки в длину начинается с разрыхления матрикса. В этом процессе важную роль играют фитогормоны. В образовавшиеся полости поступают новые порции материала, из которого строится клеточная стенка. Синтез и транспорт этих веществ осуществляются главным образом вакуолями аппарата Гольджи.
Клеточную стенку делящихся и растущих растяжением клеток называют первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои и возникает прочная вторичная клеточная стенка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений, хитин у грибов), компоненты матрикса стенки (гемицеллюлозы, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности стенки (кутин и воска). Клеточные стенки могут содержать также силикаты и карбонаты кальция.
Целлюлоза (полимер b-D-глюкозы), гемицеллюлозы (полимеры гексоз и пентоз) и пектиновые вещества (производные уроновых кислот) являются углеводными компонентами клеточных стенок. Целлюлоза и пектиновые вещества адсорбируют воду, обеспечивая оводненность клеточной стенки. Пектиновые вещества, содержащие много карбоксильных групп, связывают ионы двухвалентных металлов, которые способны обмениваться на другие катионы (Н + , К + и т.д.). Это обусловливает катионообменную способность клеточных стенок растений. Помимо углеводных компонентов в состав матрикса клеточной стенки входит также структурный белок, называемый экстенсином. Это гликопротеин, содержащий более 20% L-оксипролина от суммы аминокислот. По этому признаку белок клеточных стенок растений сходен с межклеточным белком животных - коллагеном.
Целлюлоза: А – структура молекулы целлюлозы; Б – ассоциации молекулы целлюлозы: 1 – мицелла, 2 – микрофибрилла, 3 – макрофибрилла
Основным инкрустирующим веществом клеточной стенки является лигнин. Интенсивная лигнификация клеточных стенок начинается после прекращения роста клетки. Лигнин представляет собой полимер с неразветвленной молекулой, состоящей из ароматических спиртов (п-кумарового, кониферилового, синапового). Разрушение и конденсация лигнина в почве - один из факторов образования гумуса. Интенсивная лигнификация (пропитка слоев целлюлозы лигнином) клеточных оболочек начинается после прекращения роста клетки. Лигнин может откладываться отдельными участками - в виде колец, спиралей или сетки, как это наблюдается в оболочках клеток проводящей ткани - ксилемы, или сплошным слоем, за исключением тех мест, где осуществляются контакты между соседними клетками в виде плазмодесм. Лигнин скрепляет целлюлозные волокна и действует как очень твердый и жесткий каркас, усиливающий прочность клеточных стенок на растяжение и сжатие. Он же обеспечивает клеткам дополнительную защиту от физических и химических воздействий, снижает водопроницаемость. Содержание лигнина в оболочке достигает 30%. Инкрустация им клеточных оболочек приводит к их одревеснению, которое часто влечет за собой отмирание живого содержимого клетки. Лигнин в сочетании с целлюлозой придает особые свойства древесине, которые делают ее незаменимым строительным материалом.
В регуляции водного и теплового режима растений участвуют ткани, стенки клеток которых пропитаны суберином. Отложение суберина делает стенки трудно проницаемыми для воды и растворов (например, в эндодерме, перидерме). Суберин откладывается на оболочку изнутри и делает ее практически непроницаемой для воды и растворов. В результате протопласт клетки отмирает и клетка заполняется воздухом. Такой процесс называется опробковением. Наблюдается опробковение оболочки клеток в покровных тканях многолетних древесных растений - перидерме, корке, а также в эндодерме корня. Суберин - основное вещество пробковых образований, которому эти образования главным образом обязаны своими свойствами: непроницаемостью для воды, для газов и малой теплопроводностью..
Поверхность эпидермальных клеток растений защищена гидрофобными веществами - кутином и восками: Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами стенки (целлюлозой, пектином) и образует кутикулу. Кутикула участвует в регуляции водного режима тканей и защищает клетки от повреждений и проникновения инфекции.

В оболочках эпидермальных клеток некоторых растений (злаков, осок и др.) накапливается большое количество минеральных веществ (минерализация), в первую очередь карбоната кальция и кремнезема. При минерализации листья и стебли растений становятся жесткими, твердыми и в меньшей степени поедаются животными.
В первичных клеточных стенках на долю целлюлозы приходится до 30% сухой массы стенки. Количество гемицеллюлоз и пектиновых веществ меняется в зависимости от объекта. Вместе с белками пектиновые вещества могут составлять около 30% сухой массы клетки, причем количество белка достигает 5 - 10%. Около 40% приходится на долю гемицеллюлоз.

Растительные клетки, подобно клеткам прокариот и грибов, заключены в сравнительно жесткую клеточную стенку. Материал для построения этой клеточной стенки секретирует сама заключенная в ней живая клетка (протопласт). По своему химическому составу клеточные стенки растений отличаются от клеточных стенок прокариот и грибов (табл. 2.1), но этим структурам свойственны некоторые общие функции, а именно функции опоры и защиты; кроме того, и те и другие ограничивают подвижность клеток. Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку. В этом разделе мы опишем процесс образования первичной клеточной стенки. На рис. 7.21 воспроизведена электронная микрофотография, на которой можно видеть одну из ранних стадий этого процесса.
Строение клеточной стенки
Первичная клеточная стенка состоит из целлюлозных микрофибрилл, погруженных в матрикс, в состав которого входят сложные полисахариды. Целлюлоза тоже представляет собой полисахарид (ее химическое строение описано в разд. 5.2.3). Особо важное значение для той роли, которую целлюлоза выполняет в клеточных стенках, имеют ее волокнистое строение и высокая прочность на разрыв, сравнимая с прочностью стали. Отдельные молекулы целлюлозы - это длинные полисахаридные цепи. Множество таких молекул, сшитых друг с другом поперечными водородными связями, собраны в прочные пучки, называемые микрофибриллами . Погруженные в матрикс микрофибриллы образуют каркас клеточной стенки. Матрикс клеточной стенки состоит из полисахаридов, которые для удобства описания делят обычно на пектины и гемицеллюлозы в зависимости от их растворимости в различных растворителях, употребляемых для экстракции. Пектины , или пектиновые вещества , при экстракции обычно выделяются первыми, поскольку их растворимость выше. Это - смешанная группа кислых полисахаридов (построенных из моносахаридов арабинозы и галактозы, галактуроновой кислоты, принадлежащей к классу сахарных кислот, и метанола). Длинные молекулы пектиновых веществ могут быть линейными или разветвленными. Срединная пластинка , скрепляющая стенки соседних клеток, состоит из клейких студнеобразных пектатов магния и кальция. В клеточных стенках некоторых созревающих плодов нерастворимые пектиновые вещества превращаются снова в растворимые пектины. При добавлении сахара эти последние образуют гели; поэтому их используют как желирующие вещества.
Гемицеллюлозы - это смешанная группа полисахаридов, растворимых в щелочах (к ним относятся полимеры ксилозы, галактозы, маннозы, глюкозы и глюкоманнозы). У гемицеллюлоз, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены.
Клеточные стенки гидратированы: 60-70% их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно. Присутствие воды оказывает влияние на химические и физические свойства полисахаридов клеточной стенки.
Материалы с повышенной механической прочностью, подобные материалу клеточной стенки, т. е. состоящие более чем из одного компонента, называются композиционными материалами или композитами ; их прочность обычно выше, чем у каждого из компонентов в отдельности. Системы из волокон и матрицы (в технике основу композиционного материала называют не матриксом, а матрицей. - Прим. перев) находят широкое применение в технике, так что на изучение их свойств как в технике, так и в биологии тратится много усилий. Матрица, работающая на сжатие, передает напряжение волокнам, работающим на растяжение. Она же обеспечивает абразивную стойкость и, по-видимому, стойкость к неблагоприятным химическим воздействиям, возможным в тех или иных условиях. В строительном деле издавна применяется железобетон, т. е. сочетание бетона со стальной арматурой. Позже появился более легкий композиционный материал, в котором роль матрицы играет пластик, а роль арматуры - стеклянное или углеродное волокно. Древесина представляет собой композиционный материал; своей прочностью она обязана клеточным стенкам. Примером жестких композиционных материалов биологического происхождения могут также служить кость, хрящ и покрывающая экзоскелет членистоногих кутикула. Существуют и гибкие композиционные материалы, например соединительная ткань.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной клеточной стенки (кнаружи от плазматической мембраны) отлагаются дополнительные слои целлюлозы, т. е. возникает вторичная клеточная стенка. Обычно это происходит после того, как клетка достигнет своего максимального размера, и лишь немногие клетки, например клетки колленхимы, продолжают рост во время этой фазы. Вторичное утолщение клеточных стенок растения не следует путать с вторичным утолщением (вторичным ростом) самого растения, т. е. с увеличением толщины ствола в результате добавления новых клеток.
В любом слое вторичного утолщения целлюлозные волокна располагаются под одним и тем же углом, но в разных слоях этот угол различен, чем и обеспечивается еще большая прочность структуры. Такое расположение целлюлозных волокон показано на рис. 7.27.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение); при этом все слои целлюлозы (первичный и три вторичных) пропитываются лигнином - сложным полимерным веществом, не относящимся к полисахаридам. В клетках протоксилемы отложения лигнина имеют кольцевую, спиральную или сетчатую форму, как это видно на рис. 8.11. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляются контакт между соседними клетками при помощи группы плазмодесм (разд. 8.1.3 и рис. 8.7). Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие (предотвращает прогибы). Он же обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий. Вместе с целлюлозой, остающейся в клеточных стенках, лигнин придает древесине те особые свойства, которые делают ее незаменимым строительным материалом.
Функции клеточной стенки
Ниже перечисляются основные функции клеточных стенок растений.
1. Клеточные стенки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации клеточных стенок (небольшое количество лигнина присутствует во всех клеточных стенках).
2. Относительная жесткость клеточных стенок и сопротивление растяжению обусловливают тургесцентность клеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает и в известной мере регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, окружая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт ) служит главным путем, по которому передвигаются вода и минеральные вещества. Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи, называемые плазмодесмами . Плазмодесмы связывают живое содержимое отдельных клеток - объединяют все протопласты в единую систему, в так называемый симпласт .
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой-кутикулой, состоящей из воскообразного вещества кутина, что снижает потери воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершении вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы, трахеид и ситовидных трубок (с ситовидными пластинками) приспособлены для дальнего транспорта веществ по растению. Этот вопрос рассматривается в гл. 8 и 14.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды (разд. 14.1.5).
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ; таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазматической мембраны, что повышает эффективность переноса веществ путем активного транспорта (разд. 14.8.6).
Изучен слабо. Считается, что целлюлозные микрофибриллы синтезируются на поверхности клетки с помощью ферментного комплекса, связанного с плазматической мембраной, а ориентация микрофибрилл контролируется микротрубочками , расположенными у внутренней поверхности плазматической мембраны. Пектины , гемицеллюлозы и гликопротеиды , вероятно, образуются в комплексе Гольджи и переносятся к стенке в пузырьках, отделяющихся от диктиосом .
В стенках соседних клеток, как правило, одна против другой, образуются поры .
Они чаще всего закладываются там, где есть первичные поровые поля . Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка ( рис. 22). Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной , или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется.
Содержимое соседних клеток связано друг с другом через специальные цитоплазматические тяжи - плазмодесмы . Плазмодесмы располагаются в плазмодесменных канальцах поровой мембраны . Посредством плазмодесм осуществляется передача раздражений и активное передвижение некоторых веществ от клетки к клетке.