Покровные ткани защищают растение от неблагоприятных воздействий внешней среды: солнечного перегрева, излишнего испарения, резкого перепада температуры воздуха, иссушающего ветра, механического воздействия, от проникновения вовнутрь растения болезнетворных грибов и бактерий и т.д. Как и другие постоянные ткани, покровные ткани формируются в процессе онтогенеза из меристем. Различают первичную и вторичную покровные
ткани, которые соответственно образуются в результате дифференцирования клеток первичной и вторичной меристем. Так, к первичным покровным тканям относятся кожица, или эпидерма, и эпиблема, к вторичным - перидерма (пробка, пробковый камбий и феллодерма).
Кожица, или эпидерма , покрывает все органы однолетних растений, молодые зеленые побеги многолетних древесных растений текущего вегетационного периода, надземные травянистые части растений (листья, стебли и цветки). Эпидерма чаще всего состоит из одного слоя плотно сомкнутых клеток без межклеточного пространства. Она легко снимается и представляет собой тонкую прозрачную пленку. Эпидерма - живая ткань, состоит из постенного слоя протопласта с лейкопластами и ядром, крупной вакуоли, занимающей почти всю клетку. Стенка клеток в основном целлюлозная. Наружная стенка эпидермальных клеток более утолщенная, боковые и внутренние - тонкие. Наружная стенка кожицы у злаков, осок, хвощей может быть пропитана кремнеземом, у драцен иногда встречаются кристаллы щавелевокислого кальция, у семян - полисахариды в виде слизи и т.д. На наружной поверхности клеточных стенок у некоторых растений могут откладываться суберины и кутины . Пропитанные суберином (опробковением) стенки клеток непроницаемы для воды, паров и газов. Боковые и внутренние стенки клеток имеют поры. Основная функция эпидермы - регуляция газообмена и транспирации, осуществляемая в основном через устьица. Вода и неорганические вещества проникают через поры. Эпидерма некоторых водных растений участвует в фотосинтезе, у некоторых пустынных растений в ней запасается вода.
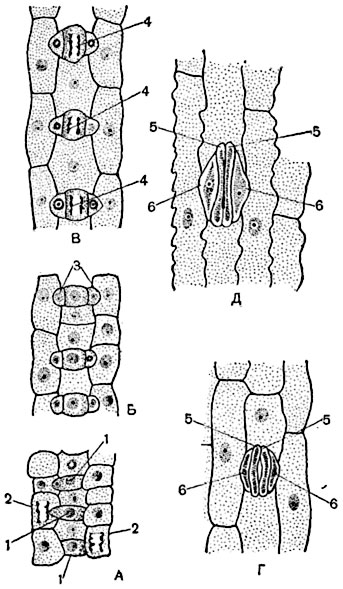
Клетки эпидермы разных растений неодинаковы по форме и размерам. У многих однодольных растений клетки вытянуты в длину, у большинства двудольных имеют извилистые боковые стенки, что повышает плотность их сцепления друг с другом (рис. 21). Эпидерма верхней и нижней частей листа также отличается своим строением: так, на нижней стороне листа в эпидерме большее число устьиц, а на верхней стороне их гораздо меньше; на листьях водных растений с плавающими на поверхности листьями (кубышка, кувшинка) устьица есть только на верхней стороне листа, а у полностью погруженных в воду растений устьица отсутствуют.
Устьица - высокоспециализированные образования эпидермы, состоят из двух замыкающих клеток и щелевидного образования между ними - устьичной щели (рис. 21, А) . Замыкающие клетки, имеющие полулунную форму, регулируют размер устьичной щели; щель может открываться и закрываться в зависимости от тургорного давления в замыкающих клетках, содержания диоксида углерода в атмосфере и других факторов. Так, днем, когда клетки устьиц участвуют в фотосинтезе, тургорное давление в устьичных
клетках высокое, устьичная щель открыта, ночью, наоборот, закрыта. Подобное явление наблюдается в засушливое время и при увядании листьев, связано с приспособлением устьиц запасать влагу внутри растения. У многих видов, произрастающих в районах с избыточным увлажнением, особенно во влажных тропических лесах, имеются устьица, через которые выделяется вода. Устьица получили название гидатоды . Вода в виде капель выделяется наружу и капает с листьев. Это происходит и с некоторыми комнатными растениями (монстера, филодендрон и другие ароидные) при снижении атмосферного давления, как правило, перед дождем. "Плач" растения - своеобразный предсказатель погоды и по-научному называется гуттацией . Гидатоды расположены по краю листа, у них нет механизма открывания и закрывания.
В эпидерме многих растений есть защитные приспособления от неблагоприятных условий: волоски, кутикула, восковой налет и др.
Волоски (трихомы) - своеобразные выросты эпидермы, они могут покрывать все растение или некоторые его части. Волоски бывают живыми и мертвыми. Волоски способствуют уменьшению испарения влаги, кроме того, они предохраняют растение от перегрева, поедания животными и от резких колебаний температуры.
Поэтому волосками чаще всего покрыты растения аридных - засушливых областей, высокогорий, приполярных районов земного шара, а также растения засоренных местообитаний.
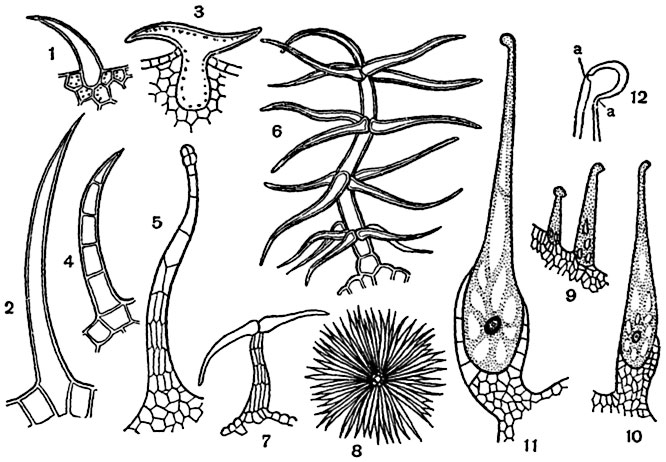
Волоски бывают одноклеточными и многоклеточными (рис. 22). Одноклеточные волоски представлены в виде сосочков. Сосочки встречаются на лепестках многих цветков, придавая им бархатистость (тагетисы, анютины глазки). Одноклеточные волоски могут быть простыми (на нижней стороне многих плодовых культур), и, как правило, они мертвые. Одноклеточные волоски могут быть ветвистые (пастушья сумка). Чаще волоски бывают многоклеточными, различающимися по строению: линейными (листья картофеля), кустисто-ветвистыми (коровяк), чешуйчатыми и звездчато-чешуйчатыми (представители семейства Лоховые), массивными (пучки волосков растений семейства Губоцветные). Встречаются железистые волоски, в которых могут накапливаться эфирные вещества (губоцветные и зонтичные растения), жгучие вещества (крапива) и др. (рис. 23). Жгучие волоски крапивы пропитаны кремнеземом, они очень ломкие. Обломившись, острые края волоска ранят кожу, на ранку выливается содержимое волоска - муравьиная кислота, которая раздражает кожу. Жгучие волоски крапивы, шипы розы, ежевики, шипы на плодах зонтичных, дурмана, каштана и др. - своеобразные выросты, называемые эмергенцами , в

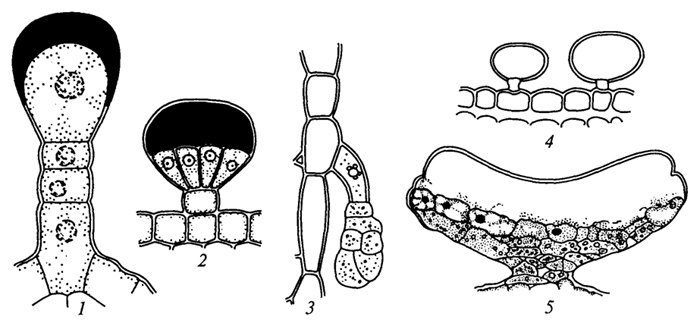
формировании которых принимают участие помимо клеток эпидермы более глубокие слои клеток.
Эпиблема (ризодерма) - первичная однослойная покровная ткань корня. Образуется из наружных клеток апикальной меристемы корня вблизи корневого чехлика. Эпиблема покрывает молодые корневые окончания. Через нее осуществляется водно-минеральное питание растения из почвы. Поскольку на корневое питание затрачивается определенная энергия, в эпиблеме много митохондрий. Клетки эпиблемы тонкостенны, с более вязкой цитоплазмой, лишены устьиц и кутикулы. Эпиблема недолговечна и постоянно обновляется за счет митотических делений.
Перидерма - сложный многослойный комплекс вторичной покровной ткани (пробка, пробковый камбий, или феллоген, и феллодерма) стеблей и корней многолетних двудольных растений и голосеменных, которые способны непрерывно утолщаться. В меньшей степени перидерма встречается у однодольных и однолетних растений. К осени первого года жизни побеги одревесневают, что заметно по изменению их окраски от зеленой до буро-серой, т.е. произошла смена эпидермы на перидерму, способную выдержать неблагоприятные условия зимнего периода. В основе перидермы лежит вторичная меристема - феллоген (пробковый камбий) , образующийся в клетках основной паренхимы, лежащей под эпидермой. Феллоген обладает слабой меристематической активностью. Он образует клетки в двух направлениях: наружу - клетки пробки , внутрь - живые клетки феллодермы , причем клеток пробки гораздо больше, чем клеток феллодермы (рис. 24). Пробка состоит из отмерших клеток, заполненных воздухом, они вытянуты в длину,

плотно прилегают друг к другу, поры отсутствуют, стенки их пропитаны суберином, клетки воздухо- и водонепроницаемы. Клетки пробки имеют коричневый или желтоватый цвет, который зависит от присутствия в клетках смолистых или дубильных веществ (пробковый дуб, бархат сахалинский). Белый цвет пробки березы обусловлен бетулином. Пробка - хороший изоляционный материал, не проводит тепла, электричества и звуки, используется для закупорки бутылок и др. Мощный слой пробки имеет пробковый дуб, виды бархата, пробковый вяз. Пробковый дуб растет в странах Средиземноморья. С плантаций пробкового дуба примерно каждые 10 лет снимают слой пробки толщиной около 10 см. На территории России в широколиственных лесах Дальнего Востока и о. Сахалин растут бархат амурский и бархат сахалинский, однако толщина пробки у них не превышает 6-7 см.
Чечевички - "вентиляционные" отверстия в пробке для обеспечения газо- и водообмена живых, более глубоко лежащих тканей растения с внешней средой. Внешне чечевички похожи на семена чечевицы, за что и получили свое название. Как правило, чечевички закладываются на смену устьицам. Формы и размеры чечевичек различны. Так, у березы чечевичка имеет узкую поперечную полоску длиной до 15 см. Однако в количественном отношении чечевичек намного меньше, чем устьиц. Чечевички представляют собой округлые тонкостенные бесхлорофилльные клетки с межклетниками, которые приподнимают кожицу и разрывают ее. Этот слой рыхлых слабо опробковевших паренхимных клеток, составляющих чечевичку, называется выполняющей тканью (рис. 25).
Корка - мощный покровный комплекс из отмерших наружных клеток перидермы. Она формируется на многолетних побегах и корнях древесных растений. У корки трещиноватая и неровная форма. Она предохраняет стволы деревьев от механических повреждений, низовых пожаров, низких температур, солнечных ожогов, проникновения болезнетворных грибов и бактерий. Растет корка за счет нарастания под ней новых слоев перидермы. У древесно-кустарниковых растений корка возникает (например, у сосны) на

8 - 10-м, а у дуба - на 25 - 30-м году жизни. Корка входит в состав коры деревьев. Снаружи она постоянно слущивается, сбрасывая с себя всевозможные споры грибов и лишайников.
Покровные ткани расположены снаружи всех органов растений на границе с внешней средой. Они состоят из плотно сомкнутых клеток и выполняют барьерную роль, предохраняя органы растений от неблагоприятных воздействий. Эти ткани возникли с выходом растений на сушу и весьма разнообразны по строению и функциям.
Покровные ткани надземных органов - эпидерма, пробка - служат для защиты от высыхания и для газообмена.
Корни первичного строения, одевает эпиблема, регулирующая и обеспечивающая всасывание и выделение растворов. Строение эпиблемы мы рассмотрим в курсе анатомии корня.
Корни вторичного строения у двудольных покрыты перидермой и коркой, а у однодольных – экзодермой.
Эпидерма (кожица). Первичная покровная ткань, образующаяся из протодермы (формирующейся из туники) конуса нарастания побега на всех листьях, стеблях, а также на цветках, плодах и семенах. Эпидерма защищает внутренние ткани от высыхания и повреждений, препятствует прониканию микроорганизмов. Одновременно эпидерма обеспечивает связь со средой - через нее происходят транспирация (регулируемое испарение) и газообмен, иногда всасывание и секреция различных веществ (в том числе эфирных масел, ферментов и гормонов).
Эпидерма - сложная ткань, в ее состав входят морфологически различные клетки: основные клетки эпидермы; замыкающие и побочные клетки устьиц; трихомы, волоски.
Основные клетки эпидермы плотно сомкнуты, межклетники отсутствуют. В эпидерме стеблей и линейных листьев они в верхней проекции продолговатые, в листьях с широкими пластинками - более или менее изодиаметрические или лопастные. Боковые стенки, т. е. перпендикулярные поверхности, часто извилистые, что повышает прочность их сцепления.
Наружные стенки обычно толще остальных. Их внутренний, наиболее мощный, слой состоит из целлюлозы и пектина. Клеточные стенки могут пропитываться кремнеземом (режущие стебли и листья хвощей, некоторых осок и злаков) или содержать слизи (эпидерма клейких, легко распространяющихся семян льна, айвы и др.).
С наружной стороны вся эпидерма покрыта сплошным слоем кутикулы. Помимо кутина в ее состав входят вкрапления воска, что еще больше снижает проницаемость кутикулы для воды и газов. На поверхности кутикулы воск может образовать сплошной налет, состоящий из чешуек, палочек и других структур. Этот сизый, легко стирающийся налет хорошо заметен на листьях капусты или плодах сливы, винограда. Если его удалить, то плоды будут быстрее портиться. Мощность кутикулы и ее состав во многом определяют химическую стойкость и проницаемость эпидермы. В условиях засушливого климата у растений развивается более толстая кутикула. У растений, погруженных в воду, кутикулы нет.
Клетки эпидермы имеют живой протопласт, обычно с хорошо развитой эндоплазматической сетью и аппаратом Гольджи. У некоторых растений (традесканции) в цитоплазме можно видеть лейкопласты. У водных растений, папоротников, обитателей тенистых мест, а также и многих светолюбивых растений (герань, подсолнечник, кактусы) встречаются хлоропласты.
Из эпидермы могут возникать придаточные почки, феллоген; в культуре можно получить зародышеподобные структуры. У некоторых, преимущественно тропических, растений эпидерма многослойна, одна из ее функций - запас воды.
Устьица - специализированные образования эпидермы, регулирующие газообмен, необходимый для дыхания и фотосинтеза, транспирации. Устьице состоит из двух замыкающих клеток, между которыми находится устьичная щель – межклетник. Под ней расположена дыхательная подустьичная полость. Она способствует лучшему газообмену между внутренними частями органа и внешней средой. Часто к замыкающим клеткам примыкают две или более побочные клетки, отличные от основных клеток эпидермы. Замыкающие и побочные клетки представляют собой устьичный аппарат.
Стенки замыкающих клеток утолщены неравномерно: брюшные (обращенные к щели) толще спинных (примыкающих к эпидерме). Замыкающие клетки содержат хлоропласты с хорошо развитыми тилакоидами и многочисленные митохондрии в активном состоянии. Устьица могут находится как в открытом состоянии так и в закрытом. Механизм открывания и закрывания устьичной щели основан на осмотических явлениях. При повышении тургора тонкие стенки растягиваются, увлекая за собой толстые, и устьичная щель увеличивается. При падении тургора она закрывается, так как замыкающие клетки принимают первоначальное положение.
Главная роль в изменении тургора и объема замыкающих клеток принадлежит ионам калия. При открывании устьиц они перемещаются из соседних клеток в замыкающие, затрачивая энергию, которую возмещают митохондрии. Существенное значение имеет и наличие хлоропластов: в результате фотосинтеза повышаются концентрация сахаров и осмотическое давление. За счет всасывания воды объем вакуоли существенно увеличивается, тургор растет и устьице открывается. В темноте при недостаточном обводнении устьичная щель закрывается из-за понижения тургора в замыкающих клетках.
Эпидерма очень эффективно регулирует транспирацию. Если устьица открыты полностью, то транспирация идет с такой же скоростью, как если бы эпидермы не было совсем. При закрытых устьицах она резко снижается.
Трихомы - различные по форме, строению и функции выросты клеток эпидермы. Они имеют форму волосков (кроющих или железистых), чешуек и др. Функции большинства типов трихом неясны. Кроющие трихомы могут быть одноклеточными (у яблони), многоклеточными неразветвленными (у картофеля) или разветвленными (у коровяка), звездчатыми (у лоха).
Волоски способны долго оставаться живыми. Но часто протопласты в них отмирают, волоски заполняются воздухом. Такие волоски защищают растение от сильной солнечной инсоляции, излишнего испарения и колебаний температуры. Многие высокогорные растения (эдельвейс) отличаются сильным опушением. Некоторые мертвые волоски, например покрывающие семена хлопчатника, достигают в длину 55 мм и широко используются в текстильной промышленности. Трихомы защищают растение от насекомых - чем гуще опушение, тем реже насекомые используют его в качестве пищи или для откладки яиц, на крючковатые трихомы насекомые и их личинки накалываются.
Помимо волосков на эпидерме образуются эмергенцы, в формировании которых принимают участие более глубоко расположенные ткани. К ним относят жгучие волоски крапивы, шипы розы, ежевики.
Эпидерма функционирует, как правило, один год, обычно к осени ее заменяет пробка.
Пробка (феллема). Вторичная покровная ткань развивается из клеток пробкового камбия, феллогена . Феллоген - вторичная меристема, он возникает из основной паренхимы, лежащей под эпидермой или более глубоко (смородина, малина), а иногда и в самой эпидерме (ива). У большинства деревьев и кустарников феллоген закладывается в однолетних побегах уже в середине лета. Клетки феллогена делятся параллельно (║) поверхности органа (тангентально), откладывая наружу клетки феллемы , внутрь - феллодермы. Клеток феллемы образуется всегда больше, чем феллодермы. Феллема (покровная ткань, пробка), феллоген (образовательная ткань) и феллодерма (основная ткань, хлорофиллоносная паренхима) - образуют единый покровный комплекс - перидерму .
Клетки пробки соединены очень плотно, без межклетников, их клеточные стенки вначале очень тонкие, затем утолщаются. Вторичные клеточные стенки состоят из слоев суберина и воска, не пропускающих воду и воздух. Опробковение стенок ведет к отмиранию протопласта. По мере того как эпидерму сменяет перидерма, зеленый цвет побегов переходит в бурый. «Вызревшие» к осени побеги первого года жизни, защищенные перидермой, способны к перезимовыванию. Пробка защищает органы растений от потери воды, проникновения болезнетворных организмов, резких колебаний температуры, так как обладает малой теплопроводностью.
Газообмен и транспирация в органах, покрытых перидермой, происходят через чечевички . Чечевичка - участок перидермы с рыхло расположенными клетками пробки. Чечевичка с поверхности выглядит как бугорок. По межклетникам этой выполняющей ткани циркулируют газы и водяные пары. С наступлением холодов феллоген откладывает под выполняющей тканью замыкающий слой из плотно соединенных клеток, препятствующий испарению. Весной этот слой под напором вновь формирующихся клеток разрывается. По мере утолщения ветвей чечевички растягиваются (у березы они имеют вид черточек, у осины - ромбов).
У древесных растений перидерма образуется на ветвях, стволах, корнях и почечных чешуях, на некоторых плодах, в местах поражения органов; у двудольных трав - на корнях, гипокотиле, иногда на корневищах, клубнях. Наиболее массивна ежегодно нарастающая пробка стволов пробкового дуба, используемая в промышленности.
Степень сформированности перидермы и ее характер необходимо учитывать при выборе способов и режима хранения овощей. В перидерме корнеплодов моркови слой пробки тонок, феллоген здесь продуцирует преимущественно феллодерму, она мощнее, чем феллема. Корнеплоды во избежание иссушения хранят в песке. Успех хранения клубней картофеля зависит во многом от сформированности перидермы. С молодых клубней слой пробки легко снимается, так как рвутся живые тонкостенные клетки феллогена. После того как феллоген дифференцируется в постоянные ткани, картофель можно закладывать на хранение.
Лишь у некоторых древесных (осины, бука, лещины) перидерма защищает стволы в течение всей жизни, а у большинства - через 10...30 лет заменяется коркой . На стволах развивается несколько перидерм, каждая последующая закладывается глубже предыдущей. Живые ткани, заключенные между слоями пробки, отмирают, и формируется покровный комплекс - корка (ритидом ). Корка состоит из нескольких слоев пробки и заключенных между ними отмерших тканей. Если образование перидерм происходит не по всей окружности ствола, а отдельными полудугами, то корка формируется неправильными кусками. Такая корка называется чешуйчатой и образуется у большинства растений. Кольцеобразная корка формируется в том случае, если каждая вновь возникающая перидерма опоясывает ствол, периодически отрезая цилиндрические участки коры (например, у винограда).
Корка не способна к растяжению, поэтому при утолщении ствола в ней появляются трещины. На дне трещин во внутренней перидерме имеются чечевички, обеспечивающие газообмен. Корка надежно предохраняет стволы от механических повреждений, лесных пожаров, резкой смены температур.
Основные ткани.
Основные ткани составляют большую часть тела растения. По происхождению основные ткани почти всегда первичны, образуются из апикальных меристем. Они состоят из живых паренхимных клеток, сильно варьирующих по форме, но в типичном случае паренхима основной ткани состоит из клеток, у которых длина немногим больше, чем ширина, чаще почти изодиаметрических. Клеточная оболочка первичная тонкостенная, с простыми порами. Иногда первичная оболочка сильно утолщается, что особенно характерно для запасающей паренхимы семян некоторых растений (Asparagus, Coffea arabica, хурма Diospyros, финиковая пальма Phoenix dactylifera). Углеводы таких оболочек рассматриваются как запасные вещества, используемые зародышем во время прорастания. Основная паренхима способна возвращаться к меристематической активности, например при заживлении ран, образовании придаточных корней и побегов.
Основные ткани выполняют функции: синтеза, накопления и использования органических веществ. В зависимости от выполняемой функции различают основную (типичную), ассимиляционную, запасающую и воздухоносную основные ткани.
Основная паренхима не имеет специфических, строго определенных функций. Она располагается внутри тела растения достаточно крупными массивами. Типичная основная паренхима заполняет сердцевину стебля, внутренние слои коры стебля и корня. Ее клетки образуют вертикальные и горизонтальные тяжи (лучи), по которым осуществляется радиальный транспорт веществ. Из основной паренхимы могут возникать вторичные меристемы.
Ассимиляционная паренхима (хлоренхима). Главная ее функция - фотосинтез. Хлоренхима расположена в надземных органах, обычно под эпидермой. Особенно хорошо развита в листьях, меньше-в молодых стеблях. Характерно наличие межклетников, облегчающих газообмен. Клетки тонкостенные, в постенном слое цитоплазмы много хлоропластов. Общий объем их может достигать 70...80 % объема протопласта.
Запасающая паренхима. Служит местом отложения избыточных в данный период питательных веществ. Запасающие ткани состоят из живых тонкостенных клеток. В крахмалоносных клетках пластиды имеют довольно простое внутреннее строение и могут быть классифицированы как амилопласты; они встречаются у многих семян и подземных запасающих органов. Паренхимные клетки в цветках и плодах часто содержат хромопласты. У некоторых семян в паренхиме запасающей ткани или зародыша хранятся твердый белок или жиры. В различных органах растения паренхимные клетки могут становиться особенно заметными благодаря накоплению в вакуолях антоцианов или таннинов или вследствие отложения в них кристаллов той или иной формы. Клетки запасающей паренхимы могут иметь толстые клеточные стенки (накапливается гемицеллюлоза в семенах финиковой пальмы), или накапливают жиры.
В этих тканях накапливаются многие растительные продукты, используемые человеком. У культурных пищевых растений обычно гипертрофированно развитие запасающей паренхимы. Запасающие ткани широко распространены, развиваются в самых разных органах. Их можно обнаружить в клубнях картофеля, корнеплодах свеклы, моркови, луковицах лука, зерновках злаков, в семенах подсолнечника, клещевины, а также в стеблях сахарного тростника, корневищах, корнях.
У растений засушливых мест - суккулентов (агавы, алоэ, кактусы) - в клетках запасающей паренхимы накапливается вода , также как у растений засоленных местообитаний (солерос). Крупные водоносные клетки есть в стеблях злаков. В вакуолях водоносных клеток имеются слизистые вещества с высокой водоудерживающей способностью.
Воздухоносная паренхима (аэренхима). Выполняет вентиляционные, отчасти дыхательные функции, обеспечивая ткани кислородом. Состоит из клеток различной формы (например, звездчатых) и крупных межклетников. Хорошо развита в органах растений, погруженных в воду (в цветоножках кувшинки, в стеблях пушицы, белокрыльника, рдеста, в корнях камыша).
Механические ткани.
Механические (опорные) ткани обеспечивают прочность растения, способность противостоять действию тяжести собственных органов, порывам ветра, дождю, снегу, вытаптыванию животными. Механические ткани имеют сильно утолщенные клеточные стенки, которые даже после отмирания протопласта продолжают выполнять опорные функцию.
У проростков, в молодых участках органов механических тканей нет, необходимую упругость они имеют благодаря тургору. По мере развития органа в нем появляются специализированные механические ткани – колленхима и склеренхима.
Колленхима.
Развивается у двудольных растений. Колленхима, которую рассматривают как опорную ткань, состоит из толстостенных клеток. Клетки колленхимы вытянуты в длину, содержат протопласты со всеми органеллами, часто содержат хлоропласты, способные к возобновлению меристематической активности. Для колленхимы характерны первичные, утолщенные и нелигнифицированные (неодревесневшие) оболочки клеток.
Наиболее характерную особенность колленхимы составляет структура клеточных оболочек. На свежих срезах оболочки выглядят толстыми и блестящими, утолщения их часто распределены неравномерно. Кроме целлюлозы они содержат большое количество пектина и гемицеллюлоз, но в них нет лигнина. Поскольку пектиновые вещества гидрофильны, оболочки клеток колленхимы богаты водой, что можно продемонстрировать, обрабатывая свежие срезы колленхимы спиртом. Обезвоживающее действие спирта вызывает заметное сжатие оболочек. На ультраструктурном уровне в оболочках клеток колленхимы различных типов обнаруживается слоистость: ламеллы с поперечной ориентацией микрофибрилл чередуются с ламеллами, в которых фибриллы ориентированы продольно. В оболочках клеток колленхимы, особенно в тех, которые имеют довольно равномерную толщину, часто встречаются первичные поры. Характер распределения утолщений клеточной оболочки изменяется в зависимости от типа колленхимы. Если оболочка утолщена неравномерно, то наибольшей толщины она достигает в углах клетки или на двух ее противоположных - наружной и внутренней - тангенциальных стенках. Колленхима с утолщениями оболочки, локализованными в углах клеток, обычно называется уголковой колленхимой , а с утолщениями на тангенциальных стенках - ламеллярной, или пластинчатой, колленхимой . С возрастом тип оболочки может меняться вследствие отложения в ней дополнительных слоев. Так, например, первоначальное распределение утолщений оболочки, характерное для уголковой колленхимы, может становиться все более неразличимым по мере того, как просвет клетки на поперечном срезе уменьшается.
Наличие межклетников не является обязательной характеристикой колленхимы. В том случае, когда межклетники присутствуют в уголковом типе колленхимы, к ним всегда обращены утолщенные стенки клеток. Колленхима с таким распределением утолщений оболочки иногда выделяется в особый тип, называемый рыхлой (лакунарной) колленхимой . Если колленхима не образует межклетников, то в углах, где соединяются друг с другом несколько клеток, срединная пластинка утолщается.
Расположение колленхимы в растении
Для колленхимы очень характерно периферическое положение. Она располагается либо непосредственно под эпидермой, либо на расстоянии одного или нескольких слоев от нее. В стеблях колленхима часто образует сплошной слой, расположенный по окружности вдоль оси стебля. Она встречается, иногда в форме тяжей, в выступающих ребрах многих травянистых стеблей и тех деревянистых, которые еще не вступили в стадию вторичного роста. В черешках колленхима распределяется так же, как и в стеблях. В листовой пластинке она находится в тех жилках, в которых проходят более крупные проводящие пучки, причем иногда она располагается по обеим сторонам жилок, а иногда только с одной, обычно нижней, стороны. Корни редко содержат колленхиму.
Связь структуры с функцией
Колленхима приспособлена, прежде всего, для выполнения функции опоры растущих листьев и стеблей. Функции опорной ткани колленхима может выполнять только в состоянии тургора. Ее оболочки начинают утолщаться на ранних этапах развития побега, однако образующееся утолщение пластично и способно к растяжению. Поэтому оно не препятствует удлинению стебля и листа. На более продвинутой стадии развития колленхима как опорная ткань сохраняется в тех частях растения (многие листья, некоторые травянистые стебли), для которых не характерна сильно развитая склеренхима. В связи с обсуждением опорной роли колленхимы интересно отметить, что в развивающихся частях растения, подвергаемых механическим нагрузкам (действию ветра или груза, приложенного к наклонным побегам), утолщение оболочки начинается раньше и становится более массивным, чем у растений, которые не подвергались таким нагрузкам.
В старых частях растения колленхима может становиться жесткой или превращаться в склеренхиму путем отложения вторичных одревесневших оболочек. Если она не претерпевает этих изменений, то ее роль как опорной ткани снижается вследствие развития склеренхимы в более глубоких частях стебля или черешка. Более того, в стеблях с вторичным ростом основной опорной тканью становится ксилема благодаря преобладанию в ней клеток с одревесневшими вторичными оболочками и обилию длинных перекрывающих друг друга клеток.
Склеренхима.
Различают первичную и вторичную склеренхиму. Первичная склеренхима развита во всех вегетативных органах однодольных, реже двудольных растений; вторичная – у подавляющего большинства двудольных. Клетки склеренхимы имеют равномерно утолщенные, как правило, одревесневшие стенки. Их прочность близка к прочности стали. Полость клетки мала, поры простые щелевидные, немногочисленные. Протопласт отмирает рано и опорную функцию выполняют мертвые клетки. Различают два основных типа склеренхимы: волокна и склереиды.
Волокна – сильно вытянутые прозенхимные клетки длиной от нескольких десятых долей миллиметра до 1 (крапива) и даже 4 см (рами). Они обеспечивают прочность органов растений на растяжение, сжатие и изгибы. Прочность волокон повышается благодаря тому, что фибриллы целлюлозы проходят в них винтообразно, меняя направление во внешних и внутренних витках. Концы клеток чаще заостренные (лен), могут быть ветвистыми (конопля), тупыми (крапива) и др.
У многих растений первичные волокна значительно длиннее, чем вторичные. Так, у конопли первичные волокна достигают 12,7 мм, а вторичные – всего 2,2 мм. Для практического использования такие особенности имеют существенное значение. Склеренхимные волокна могут встречаться в растении в виде отдельных клеток (элементарное волокно) или, соединяясь с друг другом по длине, образуют пучок (техническое волокно). Волокна выделяют с помощью мочки стебля или механически. Лучшие результаты дает мочка, когда паренхимные ткани, окружающие пучки волокон, разрушаются в результате деятельности бактерий.
Волокна стеблей двудольных растений используют для изготовления различных тканей (особо ценится неодревесневающие волокна льна) реже веревок (пенька, получаемая из конопли).
Склереиды – клетки, чаще всего имеющие паренхимную форму. Они могут располагаться в растении плотными группами или в виде одиночных клеток. Окончательно сформировавшиеся склереиды – это мертвые клетки с толстыми одревесневшими стенками, пронизанными поровыми каналами, нередко ветвистыми. Поры простые. Склереиды имеют первичное происхождение. К ним относятся каменистые (брахисклереиды) и ветвистые (астеросклереиды) клетки.
Каменистые клетки – округлые, обычно встречаются группами. Из них состоят косточки вишни, сливы, персика и скорлупа ореха. Они встречаются в сочных плодах груши, айвы, рябины и в корнях некоторых растений. В некоторых сортах груш наблюдается раздревеснение каменистых клеток при созревании плода.
Ветвистые клетки имеют разнообразную форму, играют роль опорных в листьях чая, камелии, маслины, в стеблях водных растений.
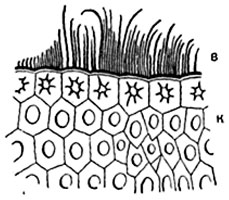
Кожица, или эпидермис . Кожица , или эпидермис , представляет собой первичную ткань листьев и стеблей. Кожица состоит из плотно сомкнутых клеток, имеющих в плане у большинства растений более или менее извилистые очертания (рис. 86). На поперечных разрезах органа клетки кожицы обычно имеют четырехугольные или пятиугольные очертания. Иногда наружная и внутренняя стенки слегка выпуклы. Клетки кожицы органов, вытянутых в длину (листовых черешков, листовых пластинок линейного и ланцетного очертания, стеблей), обычно сильно удлинены в направлении, параллельном продольной оси органа. У некоторых растений, например у многих злаков, кожица состоит из клеток нескольких типов (рис. 87).
Оболочка эпидермальных клеток утолщается обычно неравномерно: в каждой клетке наиболее толста наружная стенка, боковые стенки несколько тоньше, внутренняя - сравнительно тонка. Встречаются исключения из этого правила. Боковые и внутренние стенки клеток обычно имеют поры (рис. 87), наружная же стенка бывает снабжена ими в весьма редких случаях. Оболочка клеток состоит в основном из целлюлозы. У многих растений (особенно у злаков, осоковых, хвощей) наружная стенка кожицы пропитана кремнеземом; у некоторых растений в наружной стенке, реже в боковых (у драцен) и даже во внутренних откладываются мелкие кристаллы или зернышки щавелевокислого кальция.
Поверхность кожицы покрыта пленкой - кутикулой (надкожицей), состоящей из кутина. Кутикула очень тонка или вовсе отсутствует на поверхности конусов нарастания. Часто кутикула имеет гладкую поверхность, в иных случаях образует выступы в виде бугорков, перекладин, прямых или волнистых ребер и т. д.
Рис. 86. Участки кожицы листа лютика ползучего (Ranunculus repens ):
1 - с верхней стороны листа; 2 - с нижней его стороны.
Эти выступы состоят нацело из кутикулы или же являются выступами целлюлозной стенки, покрытыми кутикулой.
На поперечных срезах кожицы иногда видно, что кутикула пронизывает и часть толщи стенок. Кутикула сильно преломляет свет и поэтому хорошо видна на срезах. Изолированная кутикула обычно представляет бесцветную бесструктурную сплошную пленку. У некоторых растений (например, у золотого дерева Aucuba ) кутикула состоит из пластинок, каждая из которых принадлежит одной эпидермальной клетке; над пограничными участками клеток кожицы кутикулы нет или она очень тонка.
Мощность кутикулы зависит от вида и возраста растения и условий местообитания (рис.88); на поверхности стеблей с многолетней кожицей (у некоторых видов калины, клена) кутикула разрывается и подновляется изнутри путем образования новых ее наслоений. Кутикула листьев некоторых осоковых подвергается окремнению.
У многих растений во внешних стенках клеток кожицы, иногда в боковых, еще реже во внутренних, образуются кутикулярные слои - прослойки в целлюлозной клеточной оболочке, содержащие кутин (рис. 76). Кутикула и кутикулярные слои нередко пропитаны воском; при подогревании препаратов в воде под покровным стеклом он выплавляется в виде капель. У некоторых растений воск покрывает кутикулу в виде мелких зерен, расположенных равномерным слоем (на листьях и стеблях многих злаков, лилейных, например тюльпанов), тонких палочек, часто изогнутых и на конце закрученных (на стеблях некоторых злаков, сахарного тростника, рис. 89), сплошных корочек, тонких - 1? толщиной (на листьях туй, молодил).
![]()
1 - замыкающие клетки устьиц; 2 - побочные клетки; 3 - клетки эпидермиса с многочисленными порами; в полости клеток - постенная протоплазма и ядро, в некоторых из клеток не зарисованные.

Рис. 88. Часть поперечного разреза через листообразный бугорок кактуса Ariocarpus retusus
; чрезвычайно толстая кутикула пронизана фигурными ходами над устьицами. Под кожицей - зеленая паренхима.
Восковой налет, как и кутикула, снижает транспирацию органов. Воск делает поверхность органов несмачиваемой: с них быстро стекает вода, чем предупреждается капиллярное закупоривание водой устьиц и заселение поверхности растений мелкими эпифитами.
к - кожица: в - восковые палочки.
Поэтому понятно, почему стебли и листья многих подводных растений лишены кутикулы.
Клетки кожицы содержат тонкий постенный слой протоплазмы с ядром и пластидами и крупную центральную вакуолю. Обычно из пластид здесь имеются лейкопласты.
У многих растений в вакуолях клеток кожицы - нередко только на нижней стороне листа - содержится антоциан.
В вакуолях эпидермальных клеток нередко встречаются дубильные вещества, соли щавелевой кислоты в виде растворов (у щавелей, кислиц) или кристаллов; реже здесь имеются алкалоиды, ядовитые глюкозиды, гипс (у каперсов - в виде кристаллов), кальциевые соли яблочной и винно-каменной кислот (у винограда).
Хлоропласты и хромопласты в эпидермисе цветковых растений встречаются очень редко, только у некоторых теневых и водных растений .
Устьица . Для осуществления газообмена между внутренними тканями растений и внешней средой и для транспирации (испарения воды) в кожице имеются устьица.
Устьице состоит из двух специализированных замыкающих клеток и щелевидного отверстия между ними - устьичной щели . При рассматривании кусочка кожицы, положенного плашмя на предметное стекло, устьице обычно видно как пара бобовидных или полулунных клеток, обращенных друг к другу вогнутыми сторонами (рис. 90-92). Оболочка каждой замыкающей клетки на внутренней стороне сильно и неравномерно утолщена, на противоположной, наружной, стороне - тонка.
На поперечном разрезе, проведенном через середину устьичной щели перпендикулярно к ней (рис. 91), видны выросты стенок замыкающих клеток устьица, называемые отрогами или клювиками.
Щель устьица имеет сложное очертание: промежуток между наружными отрогами ведет в преддверие, или передний дворик устьица . Дальше идет наиболее узкая часть устьичной щели - центральная щель устьица , находящаяся между тонкими участками внутренних стенок. Центральная щель расширяется дальше в задний дворик . Отверстие между отрогами нижних стенок замыкающих клеток
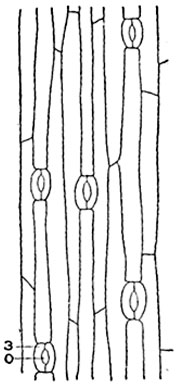
Рис. 90. Кожица листа гиацинта (Hyacinthus orientalis
) с устьицами, в плане:
з - замыкающие клетки, о - щель устьица.

Рис. 91. Устьице с окружающими клетками эпидермиса (листья тимьяна Thymus ):
1 - в плане; 2 - в разрезе; щ - центральная щель устьица; а - тонкая стенка замыкающей клетки устьица, обращенная к соседним клеткам кожицы; в замыкающих клетках видно ядро (я ) с ядрышком и хлорофилловые зерна (x ); к - кутикула; м - межклетная воздухоносная полость ("дыхательная полость").

Рис. 92. Устьичный аппарат листа свеклы (Beta vulgaris
):
1, 3 - устьице в плане; 2, 4 - в разрезе; 1, 2 - устьице закрыто; 3, 4 - устьице открыто.
сообщает задний дворик с дыхательной полостью (рис. 91, 2 ; 93).
Возвращаясь к устьицу в плане (рис. 91, 1 ), отметим, что щель устьица в оптических сечениях, параллельных поверхности эпидермиса, имеет форму узкой двояковыпуклой линзы; в плане обычно видно очертание щели на уровне наружного отверстия и на уровне середины центральной щели. Наружные стенки, а у некоторых растений отчасти также и внутренние покрыты кутикулой; оторочки двориков иногда состоят сплошь из кутикулы. Функционирующие замыкающие клетки живые; они содержат постенный слой протоплазмы с ядром и с хлорофилловыми зернами.
Обычно устьица распределяются на поверхности листа или стебля поодиночке и более или менее равномерно (рис. 90). У некоторых растений (например, у злаков) они располагаются продольными рядами, а у других (у олеандра, селезеночника) - группами.
Образование замыкающих клеток устьиц происходит при неравномерном делении меристематических клеток эпидермиса, в результате чего возникают инициальные клетки устьиц. В простейших случаях инициальная клетка становится производящей клеткой устьица.
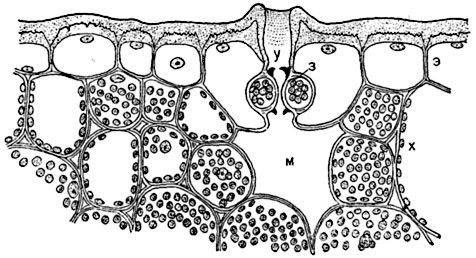
Рис. 93. Часть поперечного разреза листа Aloe disticha с устьицем, погруженным в ткань листа:
э - кожица с толстыми наружными стенками, покрытыми мощной кутикулой; у - полость над устьицем; з - замыкающие клетки с кутикулярными выступами (обозначенными черным цветом); м - воздухоносный межклетник ("дыхательная полость"); х - клетки хлорофиллоносной паренхимы.
Производящая клетка образует путем деления пару замыкающих клеток устьица; расщепление перегородки между ними ведет к образованию устьичной щели. У некоторых растений инициальная клетка образует путем деления целые группы клеток - пару замыкающих и две или несколько побочных клеток устьица; последние отличаются от прочих клеток кожицы меньшей величиной и своеобразными очертаниями (рис. 94).
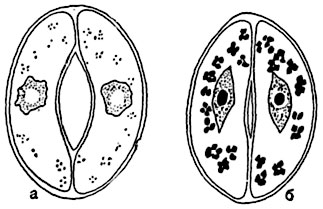
Устьичный аппарат регулирует газообмен растения. Замыкающие клетки могут смещаться, изменять объем и форму, в результате чего меняется и очертание устьичной щели: она может быть более или менее широко открыта или же вовсе закрыта (рис. 92). При открывании и закрывании устьиц происходят изменения живого содержимого замыкающих клеток. При открытой щели устьица протоплазма в них более вязка, чем при закрытой. Ядро изменяет форму: у кормовых бобов при длительно открытом устьице оно становится веретеновидньш, при закрытом - округло-лопастным, у георгина - наоборот (рис. 95).
Открывание и закрывание устьиц обусловливается тургорными явлениями. Изменения тургора замыкающих клеток могут быть непосредственным следствием подвядания листа или повышения в нем содержания воды.
При повышении тургора замыкающих клеток объем их клеточной полости увеличивается, в результате чего стенки отдаляются одна от другой и щель открывается. При понижении тургора замыкающих клеток происходят изменения обратного рода: стенки сближаются до соприкосновения и щель устьица замыкается.
Изменения тургора в замыкающих клетках регулируются превращением в них крахмала хлоропластов в сахар, и обратно - сахара в крахмал: при осахаривании крахмала в замыкающих клетках повышается концентрация клеточного сока и увеличивается их сосущая сила. В результате замыкающие клетки насасывают воду (частично за счет соседних клеток), что вызывает изменение их объема и раскрывание устьичной щели (рис. 96). Превращение сахара замыкающих клеток в крахмал влечет аналогичным образом закрывание щели устьица.
Косвенное участие в закрывании и открывании устьиц могут принимать не только клетки, соседние с замыкающими, но и более дальние побочные клетки устьиц и рядовые клетки кожицы. При продолжительном дожде все клетки кожицы насыщаются водой, тургор их сильно повышается и замыкающие клетки устьиц сдавливаются; в результате устьичные щели пассивно замыкаются. Через некоторое время после прекращения дождя нормальная работа устьиц восстанавливается.
Тургорная теория действия устьиц, выдвинутая Молем в 1856 г., подверглась дополнениям со стороны ряда ученых, указывающих на возможное значение изменений проницаемости протоплазмы замыкающих клеток.
В устройстве устьиц встречается большое разнообразие. Форма щели может быть значительно упрощена или усложнена по сравнению с вышеописанной, типичной. Наиболее упрощена она у растений с листьями, плавающими на воде (у белой кувшинки): щель имеет вид воронки, широким концом соединяющейся с дыхательной полостью, а узким переходящей в наружное отверстие; степень раскрытия щели устьица зависит от состояния этого отверстия.
У растений с листьями, расположенными более или менее горизонтально, особенно же у деревьев и у трав тенистых местообитаний, устьица расположены преимущественно или исключительно на их нижней стороне.
У многих растений, преимущественно у приуроченных к сухим, солнечным и ветреным местообитаниям, устьица погружены в глубь органа за счет сильно кутинизированных выступов соседних клеток (рис. 93). Иногда устьица располагаются целыми группами в глубине ямочек (как у олеандра). У некоторых видов австралийских бэнксий (Banksia ) на дне и вокруг ямок с устьицами имеются длинные белые волоски; образуются, таким образом, убежища с затишьем от ветра и затенением от солнца.
А - три производящие клетки устьиц (1, 1, 1 ) уже отделились перегородками от инициальных клеток; в соседних клетках происходит деление ядра (2, 2 ), предшествующее образованию побочной клетки; Б - при каждой из производящих клеток трех устьиц образовались две побочные клетки (3 ); В - в трех производящих клетках устьиц происходит деление ядра (4, 4 ), предшествующее образованию замыкающих клеток; Г - производящая клетка поделилась на две клетки, дифференцирующиеся в пару замыкающих клеток устьица (5, 5 ), с устьичной щелью между ними; 6, 6 - побочные клетки; Д - сформированный устьичный аппарат, состоящий из двух замыкающих клеток (5, 5 ) и двух побочных клеток (6, 6 ).
Волоски (трихомы ). У большинства высших растений некоторые или многие из клеток кожицы образуют выросты, так называемые волоски , имеющие различную форму, сосочков, бугорков, щетинок, кустиков, звездчатых чешуек и т. д. В простейшем случае волосок представляет сосочек - короткий округло-конический вырост клетки кожицы.
а - устьице открыто, в замыкающих клетках очень мелкие крахмальные зерна, ядро округло-амебоидного очертания, с вакуолей в центре; б - устьице закрыто, в замыкающих клетках - сравнительно крупные зерна крахмала, клеточное ядро веретенообразно, с крупными ядрышками в центре.
Сосочками же являются волоски почти всех типов в ранней стадии их образования. Встречаются волоски, не отделяющиеся перегородкой от произведшей их клетки кожицы, как например щетинки многих бурачниковых. В большинстве случаев волосок отделяется перегородкой от произведшей его эпидермальной клетки - "материнской клетки волоска". Отчленяющийся волосок редко остается одноклеточным; чаще он становится многоклеточным. Многоклеточные волоски очень разнообразны. Среди них различают 1) линейные, состоящие из клеток, расположенных в ряд (листья картофеля); 2) кустисто-ветвистые (коровяки, рис. 97, 6 ); 3) чешуйчатые и звездчато-чешуйчатые (Elaeagnus , рис. 97, 7, 8 ) и 4) массивные, представляющие как бы пучок спаявшихся нитевидных волосков (волоски многих губоцветных). Содержимое волосков некоторых категорий во взрослой стадии разрушается, полости их обычно заполняются воздухом. Такие волоски, имеющие обычно белый, либо сероватый, или желтоватый цвет, можно считать одним из приспособлений, умеряющих действие на лист или стебель горячих лучей солнца и ослабляющих иссушающее действие ветров.
Некоторые волоски могут служить растению защитой от поедания животными; так, тесно расположенные жесткие, острые щетинки и войлочный покров предохраняют листья и стебли от объедания слизняками и отчасти от поедания травоядными млекопитающими.
Жгучие волоски . Жгучий волосок крапивы - крупная живая клетка с пузыревидным основанием, сидящим в чашевидной многоклеточной "подставке". Волосок представляет в миниатюре как бы полую иглу шприца и заканчивается косо поставленной головкой. Так как оболочка волоска, богатая известью и кремнеземом, очень ломка и к тому же в месте перехода "иглы" в "головку" весьма тонка, то при малейшем прикосновении животного или человека к головке последняя отламывается, острый край "иглы" вонзается в кожу, и клеточный сок волоска, содержащий жгучие вещества, впрыскивается в нее (рис. 97, 9-12 ).
Прикосновение руки к листьям некоторых крапив тропической флоры (например, Urtica urentissima Индонезии) вызывает болезненное или даже опасное для жизни воспаление.
К покровным тканям относятся еще перидерма и корка (описание их см. на стр. 173-175).
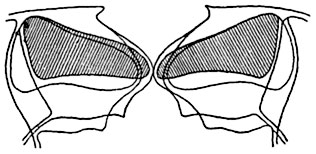
Рис. 96. Схематизированный оптический разрез устьица зимовника (Heiieborus sp
.) в закрытом и открытом состоянии.
Наружные очертания и клеточная полость замыкающих клеток показаны при закрытой щели устьица и при открытой щели. Клеточная полость обеих замыкающих клеток при закрытом устьице заштрихована.
Покровные ткани
Покровные ткани растений. Покровные ткани. Основные функции - защита молодых органов от высыхания, механическая защита и газообмен. Различают: эпидерму, перидерму и корку. 1. Эпидерма, первичная покровная ткань. Образована одним слоем клеток, покрывающих все молодые органы растений. Покровная ткань зоны всасывания корней называется эпиблемой (ризодермой).
Покровные ткани являются пограничными. Они располагаются на поверхности органов растений, что обусловливает особенности их строения и многообразие выполняемых функций.
1. Покровные ткани обеспечивают связь растений с окружающей средой. При этом они являются не просто пассивным барьером, но и активными участниками взаимодействий. Например, клетки эпидермы участвуют в определении совместимости пыльцы и рыльца пестика; в развитии процессов, связанных с иммунитетом растений, реакцией на изменение температуры и влажности воздуха и почвы.
2. Покровные ткани осуществляют транспирацию и газообмен и их регулирование.
3. Покровные ткани участвуют в выделении продуктов метаболизма.
4. Покровные ткани защищают растения от поражения болезнетворными вирусами, бактериями, грибами; от поедания животными.
5. Покровные ткани предохраняют растения от суточных и сезонных перепадов температуры.
С учётом происхождения, местоположения, особенностей клеточного строения и выполняемых функций среди покровных тканей выделяют эпидерму, эпиблему, веламен, перидерму и корку. По мнению ряда учёных сюда же относится и семенная кожура.
Первичные покровные ткани. Эпидермис. Эпиблема. Веламен
Эпидерма (эпидермис) является первичной покровной тканью, характерной для всех высших растений. У покрытосеменных она располагается на поверхности листьев, стеблей и их видоизменений, а также на репродуктивных органах – цветках и плодах. Кроме того, эпидермальные клетки могут выстилать внутреннюю поверхность эндокарпия – внутреннего слоя плодовой оболочки. Эпидерма характерна для травянистых растений. У деревьев и кустарников она развивается на молодых частях побегов, а по мере их старения замещается перидермой.
Эпидерма возникает из протодермы, порождаемой туникой конуса нарастания побега. Это сложная ткань, в состав которой входят основные и побочные клетки, замыкающие клетки устьиц, а также трихомы – выросты эпидермы. Клетки эпидермы располагаются однослойно. Однако у некоторых тропических растений (пальмы, орхидеи и др.) эпидерма бывает многослойной.
Основные клетки эпидермы стеблей, черешков листьев, а у однодольных и листовых пластинок, имеют удлиненную форму, вытянутую вдоль оси органа. Например, в стеблях злаков длина клеток эпидермы достигает 300 мкм. В листовых пластинках большинства двудольных, на цветках и плодах эпидермальные клетки имеют паренхимную и складчатую форму. Основные клетки эпидермы содержат развитые вакуоли, но лишены хлоропластов и не принимают участия в фотосинтезе. Их клеточные оболочки имеют ряд особенностей. Внутренняя оболочка тонкая, гладкая. Антиклинальная, т.е. поперечная, часто бывает извилистой, что увеличивает площадь соприкосновения смежных клеток эпидермы, а следовательно – силу их сцепления. Наружная оболочка наиболее прочная. Она образована более толстым слоем целлюлозы. Химический состав оболочек изменяется в ходе онтогенеза. У хвощей, осок и злаков оболочки часто пропитываются солями кальция, у овса – кремнеземом, у сахарного тростника – суберином. Реже, у бамбуковидных мятликовых, наблюдается одревеснение оболочек. В цитоплазме эпидермальных клеток могут образовываться кристаллы: рафиды (виноград, недотрога), друзы (свекла), цистолиты (фикус), реже – кристаллический песок (картофель).
Протопласт эпидермы активно вырабатывает и выделяет на наружную поверхность высокомолекулярные органические соединения, которые в совокупности образуют кутикулу. В составе катикулы первый слой образуют разветвленные молекулы пектина, затем располагаются глыбки растительного воска, кутин и эпикутикулярный воск. В зависимости от плотности сложения и скульптурированности поверхностного слоя, кутикула может быть плотной и блестящей, как у листьев фикуса, либо рыхлой и матовой, как у стеблей ржи, плодов сливы или яблони. Кутикула снижает интенсивность транспирации, отражает лучи солнца и защищает лист от перегрева, уменьшает возможность проникновения патогенов в растение.
Среди основных клеток эпидермы на верхней стороне листа мятликовых встречаются крупные моторные клетки с большими вакуолями. В жаркую пору они обеспечивают сворачивание листа в трубку (кукуруза) или складывание листовой пластинки (ковыль), что защищает растение от избыточного испарения влаги.
На основных клетках эпидермы часто образуются выросты, или трихомы, которые бывают по выполняемым функциям кроющими и железистыми, а по структуре – волосками и эмергенцами. Кроющие волоски заполнены воздухом и имеют белёсую окраску. Они хорошо отражают прямые солнечные лучи и уменьшают испарение воды. Железистые волоски – это живые клетки, вырабатывающие и выделяющие эфирные масла и другие конечные продукты метаболизма. Волоски образуются из клеток эпидермы. Они бывают одноклеточными и многоклеточными, простыми и ветвистыми. Одноклеточные простые волоски характерны для термопсиса и подмаренника, одноклеточные разветвленные – для пастушьей сумки и маттиолы. Многоклеточные простые встречаются у пеларгонии, многоклеточные ветвистые – у лаванды, а звездчатые – у лоха и облепихи.
Эмергенцы – это поверхностные структуры, образуемые клетками эпидермы и более глубоко расположенных тканей. К ним относятся шипы на плодах у сельдерейных, шипы на стеблях малины и ежевики, жгучие волоски листьев и стеблей крапивы. Они защищают растения от поедания.
Замыкающие клетки устьиц располагаются между основными клетками эпидермы. Они являются активными участниками транспирации и газообмена. Замыкающие клетки имеют небольшие размеры, содержат хлоропласты и попарно располагаются в эпидерме, не срастаясь между собой. Между ними находится устьичная щель, а под ними – подустьичная воздушная полость.
У однодольных замыкающие клетки имеют гантелевидную форму. Они сужены в средней части и расширены по краям. В расширенной части клеточные оболочки тонкие и растяжимые, а в середине – утолщенные и нерастяжимые. У двудольных замыкающие клетки имеют, при просмотре сверху, бобовидную форму. У них утолщены и нерастяжимы участки оболочки, обращенные к устьичной щели. Наружные оболочки этих клеток имеют выросты, которые возвышаются над устьичной щелью и образуют небольшую полость – передний дворик. Если такие же выросты возникают ниже устьичной щели, то они образуют задний, или внутренний, дворик, который непосредственно связан с подустьичной воздушной полостью. В совокупности замыкающие клетки, устьичная щель, передний и задний дворик, а также подустьичная воздушная полость образуют устьице, регулирующее транспирацию и газообмен. Число устьиц достаточно велико. У культурных растений на 1 мм 2 листовой поверхности размещается от 100 до 700 устьиц.
Работа устьиц сводится к регулированию ширины устьичной щели посредством сложных физиологических процессов. Устьица могут открываться в разное время суток. Это зависит от физиологического состояния растений, химического состава и концентрации клеточного сока, обеспеченности клеток водой и углекислым газом, освещенности листа, температуры воздуха и силы ветра.
Утреннее открывание устьиц можно схематично представить следующим образом. Под влиянием света в хлоропластах замыкающих клеток проходит фотосинтез. В результате концентрация их клеточного сока увеличивается, что приводит к поступлению воды из соседних, побочных, клеток и возрастанию тургорного давления. Под действием тургорного давления тонкие участки оболочек замыкающих клеток растягиваются в стороны, противоположные от устьичной щели, и толстые участки оболочки отходят друг от друга. Устьице открывается. При снижении тургорного давления в замыкающих клетках устьица закрываются. Подробно работа устьиц и процесс транспирации изучаются в курсе физиологии растений.
Побочные клетки располагаются рядом с замыкающими клетками устьиц и участвуют в регулировании их водного режима. По размерам они меньше основных и не имеют хлоропластов. Расположение побочных клеток является диагностическим признаком, используемым в систематике растений. Известно более десятка типов устьиц в зависимости от взаимного расположения замыкающих и побочных клеток. Например, при аномоцитном типе, характерном для винограда, красного перца и других цветковых растений, многочисленные побочные клетки не отличаются по размерам от основных клеток эпидермы. При перицитном, известном у папоротников – одна побочная клетка окружает устьице. У двудольных часто встречается парацитный тип устьица, когда две небольшие побочные клетки располагаются параллельно замыкающим. Для однодольных характерен тетрацитный тип, где четыре побочные клетки расположены вокруг устьица так, что плоскости их стенок не совпадают с плоскостью расположения устьичной щели.
Расположение устьиц на поверхности листа также имеет систематическое значение. Например, у однодольных они располагаются ровными рядами, а у двудольных – рассеяно по всей поверхности листовой пластинки. Как правило, устьица располагаются на нижней стороне листа. Это уменьшает испарение воды наземными растениями с плагиотропным, т.е. параллельным поверхности Земли расположением листьев (яблоня, свекла и др.). Если лист тяготеет к вертикальному, ортотропному, положению в пространстве, то устьица на нем могут располагаться с обоих сторон, как у кукурузы или лука. У водных растений, листья которых плавают на поверхности воды, устьица находятся на верхней стороне, как у типичной для белорусских водоёмов кубышки желтой.
Эпиблема. Особым видом первичной покровной ткани является эпиблема, которая образуется меристематическими клетками поверхности кончика корня. Клетки эпиблемы живые, тонкостенные, с развитой вакуолью, располагаются на поверхности корня в зоне всасывания. Одни из них, трихобласты, образуют особые одноклеточные выросты – корневые волоски, которые активно всасывают из почвы воду и растворенные в ней вещества. Другие, атрихобласты, корневых волосков не имеют. Жизнь корневых волосков ограничена двумя – тремя неделями. Затем они разрушаются и слущиваются. Одновременно, по мере роста кончика корня, на нем образуются новые корневые волоски.
Эпиблема на корнях однодольных сохранается на протяжении всей жизни растения. У двудольных в связи с переходом корня к вторичному анатомическому строению эпиблема сохраняется лишь в зоне корневых волосков, а в зоне проведения она замещается перидермой.
Веламен – это первичная покровная ткань воздушных корней тропических эпифитов и лазающих лиан.
Эпифиты произрастают в щелях ветвей, заполненных гумусом, и у оснований листьев крупных деревьев, используя их лишь как опору или субстрат, например, тениофиллум и фаленопсис из семейства Орхидные. Они поселяются также на скалах. Лазающие лианы, к которым относятся культурные виды – ваниль плосколистная (сем. Орхидные) и монстера деликатесная (сем.Ароидные), в качестве субстрата используют почву. У них образуются длинные стебли, прикрепляющиеся к опорным деревьям с помощью воздушных корней, что позволяет выносить листья и цветки в верхний ярус, к свету. У многих из них, как у монстеры, на стебле образуются питающие воздушные корни, растущие вниз. Достигнув почвы, они проникают в неё, ветвятся и всасывают воду и минеральные вещества.
Веламен образуется из протодермы и состоит из нескольких слоев мертвых клеток, оболочки которых имеют сетчатые или спиральные утолщения, придающие им прочность. Благодаря наличию в оболочках многочисленных пор и сквозных отверстий, веламен как губка капиллярным путём всасывает дождевую воду, а также конденсирующуюся атмосферную влагу. Таким образом, веламен выполняет защитную роль и регулирует водный режим растений.
Вторичный покровный комплекс – перидерма
Перидерма является вторичным покровным комплексом, который характерен для голосеменных и двудольных покрытосеменных. Она образуется на корнях и корневищах, клубнях и стеблях, на листовых рубцах и кроющих чешуях зимующих почек, реже – на плодах. Раневая перидерма формируется на травмированных участках органов растений.
Перидерма порождается клетками пробкового камбия, именуемого феллогеном. Феллоген имеет все признаки образовательной ткани. Это небольшие прямоугольные, слегка утолщенные в радиальном направлении, плотно сложенные клетки с тонкими оболочками. Они имеют густую вязкую цитоплазму и многочисленные органоиды, связанные с процессами синтеза, обеспечивающими деление ядра и цитоплазмы.
Феллоген образуется, как правило, в первый год жизни органа, например, на побегах яблони или смородины. Однако у тисса ягодного он появляется на 2 – 3 год, а у клена американского – на 3 – 4 год. Феллоген закладывается в поверхностных слоях растений. Например, у розы он возникает в эпидерме, у малины – в первичной коре, у спиреи – в перицикле, у пузыреплодника – во вторичной флоэме. При формировании вторичного анатомического строения корня феллоген образуется из клеток перицикла, а при образовании корки повторные слои феллогена возникают во вторичной коре.
Митотическое деление клеток феллогена приводит к образованию феллемы и феллодермы. Феллема откладывается наружу от феллогена. Её клетки располагаются многослойно, ровными рядами, одни над другими. У разных групп растений клетки феллемы отличаются толщиной и видоизменениями оболочек, наличием кристаллов оксалата кальция. При пропитывании оболочек феллемы суберином образуется пробка. Возможно также образование феллоида, т.е. ткани, которая появляется при пропитывании оболочек феллемы лигнином.
Мертвые клетки пробки имеют плотное сложение, они заполнены воздухом, либо загустевшей протоплазмой темного цвета. Пробка непроницаема для воды и имеет низкую теплопроводность, поэтому эффективно защищает растения от потери воды, перепадов температуры, болезнетворных микроорганизмов. Развитие пробки оказывает прямое влияние на сохранность клубней картофеля и корнеплодов в период роста и хранения. Пробка пробкового дуба имеет большое значение в производстве качественных вин. Феллодерма, образуемая феллогеном, является живой паренхимной тканью. Её клетки содержат хлоропласты и способны накапливать крахмал. Пробка в совокупности с феллогеном и феллодермой составляет вторичный покровный комплекс – перидерму. В отдельных местах перидермы образуются чечевички, которые служат для газообмена и регулирования водного режима растений. Чечевички – это отверстия в пробке, заполненные выполняющей тканью, состоящей их рыхло сложенных, слабо опробковевших паренхимных клеток. Под чечевичкой в феллогене имеются небольшие межклетники, облегчающие газообмен живых клеток феллодермы. В конце лета под выполняющей тканью чечевички феллоген производит слой мелких, плотно сложенных клеток замыкающего слоя, который значительно уменьшает газообмен и испарение воды через чечевички в период сезонного покоя растений. Весной замыкающий слой разрушается под давлением вновь образуемых клеток пробки и чечевички продолжают выполнять свои функции. Продолжительность сохранности перидермы зависит от видовых особенностей растений. Наиболее коротка она у травянистых растений. У садовых кустарников (смородина, крыжовник) перидерма сохраняется всю жизнь, у плодовых семечковых пород (яблоня, груша) – в течение 5 – 10 лет. Бук, граб, черёмуха, лещина на протяжении всей жизни остаются гладкоствольными.
Корка
У большинства деревьев на смену перидерме образуется корка или ритидом. Корка возникает из поверхностных слоев коры в результате заложения повторных перидерм. Феллоген этих перидерм формируется из паренхимы вторичной флоэмы. При этом живые паренхимные клетки коры, изолированные слоями пробки, отмирают и в совокупности с другими тканями коры и слоями повторной пробки образуют корку. Если повторные перидермы образуются в виде неправильных дуг, которые своими концами соприкасаются с ранее возникшими перидермами, то образуется трещиноватая, или чешуйчатая, корка (дуб, клён и др.). При заложении повторной перидермы сплошным кольцом, как у винограда, корка будет кольчатой. При утолщении ствола мёртвые ткани корки растрескиваются в отдельных местах, однако трещины не доходят до живых тканей коры. Темпы нарастания корки в толщину невелики, так как её наружные слои постепенно слущиваются. Корка выполняет защитную функцию, предохраняя стволы деревьев от механических повреждений, перепадов температуры, проникновения патогенов. Однако в её трещинах могут сохраняться зимующие стадии вредоносных насекомых, споры болезнетворных грибов и бактерий. Это обусловливает необходимость постоянного ухода за поверхностью стволов древесных садовых пород.
Семенная кожура
Особым типом покровной ткани является семенная кожура или семенная оболочка, которая образуется из покровов (интегументов) семязачатка и является обязательной частью семени. Она состоит из нескольких слоёв плотно сложенных клеток и выполняет защитную роль, предохраняя зародыш семени и запасающие ткани от пересыхания, механических повреждений и болезнетворных микроорганизмов. Ослизнение семенных оболочек, как у бобовых, льна и других растений, способствует всасыванию влаги из почвы. Кроме того, прочная кожура может защищать семена от преждевременного прорастания, что в агрономической практике называется твердокаменностью. В связи с этим семена некоторых культур (клевер, галега и др.) скарифицируют перед посевом, т.е. разрушают их семенные оболочки либо путем механического перетирания с песком и другими материалами, либо воздействием серной или других кислот. Выросты семенной оболочки в виде волосков опушения, как у ив и хлопчатника, обеспечивают распространение семян ветром. Толщина кожуры часто связана с развитием плодовой оболочки. У растений с прочным невскрывающимся околоплодником она, как правило, более тонкая, например у орехоплодных. Важными структурными образованиями на поверхности кожуры являются микропиле и рубчик. Микропиле – это небольшое отверстие, через которое начинается проникновение воды при прорастании семени. Рубчик является местом прикрепления семени к семяножке.
Располагаются на границе с внешней средой. Большинство состоит из плотно сомкнутых живых, реже мертвых клеток.
Выполняют барьерную роль, защищая внутренние органы от высыхания и повреждения.
Покровные ткани - барьер для проникновения патогенных микроорганизмов. Образовалась она в процессе эволюции в момент выхода растений из водной среды на сушу. Она возникает из меристем.
Какие первичные покровные ткани различают?
Различают первичные покровные ткани:
1. Первичные - эпидерма и эпиблема
2. Вторичные - перидерма (пробка), образуется из феллогена
3. Третичная - ритидом или корка.
Эпидерма и ее основные особенности:
Эпидерма : листья и молодые побеги как мехом покрытые однородной первичной покровной тканью - эпидермой. Она возникает из конуса нарастания туники. Наружная поверхность клеток эпидермы часто покрыта слоем кутикулы. Она может достигать значительной толщины.
Отсутствуют межклетники, клетки плотно сомкнуты. Главная функция эпидермы регуляция газообмена и транспирация, т.е. испарение воды растением. Они осуществляются через устьица, но могут частично и через кутикулу. Форма клеток эпидермы различна. Внутри клетки присутствует одна крупная вакуоль.
Обычно клетки эпидермы бесцветны, но иногда, особенно в клетках плодов цветков, могут быть окрашенными. У некоторых растений под эпидермой расположена особая ткань - гиподерма (у хвои сосны).
Выполняет механическую функцию и предохраняет от испарения.
Производные эпидермы:
Устьица - высоко специализированные образования эпидермы состоят из двух замыкающих клеток и устьичной щели. Стенки замыкающих клеток утолщены не равномерно.
Брюшные (около щели) толще сменных. Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под ней лежит дыхательная или воздушная полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим называются побочные или около устьичными. Вместе они образуют устьичный аппарат.
От строения устьичного аппарата зависит устьичный тип. Их изучение получило название стоматография («стома» - от греч. устьице). Данные могут использоваться в систематике растений и в фармакогнозии для микродиагностики лекарственного растительного сырья.
Устъичные типы:
1. Аномоцитный тип - (аномос - беспорядочный). Побочные клетки не отличаются от остальных эпидермальных клеток, характерны для всех групп высших растений, исключая хвойные.
2. Диацитный тип - есть только две побочные клетки, общая стенка которых находится под прямым углом к замыкающим клеткам (губоцвентные гвоздичные).
3. Парацитный тип - (пара - рядом). Пробочные клетки расположены параллельно замыкающим и устьичной щели (папоротники, хвощи, ряд цветковых).
4. Анизоцитный тип - (анизос - неравный) замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных (только у цветковых растений).
5. Тетрацитный тип - (тетра - четыре) замыкающие клетки окружены четырьмя побочными клетками (однодольные).
6. Энциклоцитный тип - (циклос - колесо). Побочные клетки образуют узкое кольцо вокруг замыкающих клеток (папоротники).
7. Актиноцитный тип - (актис - луч). Побочные клетки радиально расходятся от замыкающих клеток. Этот тип клеток встречается только у цветковых растений.
Волоски, выстилающие устьица, называются устьичными криптами. Число устьиц на листе сильно варьирует от 10-20 до 200-300 на 1 кв.мм. Механизм их работы очень сложен и зависит от температуры, света, воды. Они составляют 1-2% от площади листа.
Что такое эпидермальные трихомы?
Волоски на эпидерме называются трихомы . Они делятся на кроющие и железистые. Железистые - это производные секреторных тканей. Кроющие обычно расположены на той же стороне, где расположены устьица. Трихомы - различные по форме, строению и функции выросты клеток эпидермы.
Формы эпидермальных трихом:
Они имеют форму волосков (кроющих или железистых, которые будут рассмотрены в составе выделительных тканей), чешуек и др. Функции большинства типов трихом неясны. Кроющие трихомы могут быть одноклеточными (у яблони), многоклеточными неразветвленными (у картофеля) или разветвленными (у коровяка), звездчатыми (у лоха)
Немного про волоски …
Волоски способны долго оставаться живыми. Но часто протопласты в них отмирают, волоски заполняются воздухом. Такие волоски защищают растение от сильной солнечной инсоляции, излишнего испарения и колебаний температуры.
Многие высокогорные растения (эдельвейс) отличаются сильным опушением. Некоторые мертвые волоски, например, покрывающие семена хлопчатника, достигают в длину 55 мм и широко используются в текстильной промышленности. Трихомы защищают растение от насекомых
Чем гуще опушение, тем реже насекомые используют его в качестве пищи или для откладки яиц, на крючковатые трихомы насекомые и их личинки накалываются.
Выросты на эпидерме называются эмергенцы - это жгучие волоски крапивы, шипы розы, малины, ежевики, шипы на плодах дурмана, каштана.
Эпидерма функционирует, как правило, один год, обычно к осени ее заменяет пробка.
Эпиблема: не редко называется ризодерма. Она возникает из дерматогена, через нее идет поглощение воды и минеральных солей из почвы. Это волосконосный слой во всасывающей зоне коры. Корневые волоски формируются не у всех клеток эпидермы коры, а через специальные трихобласты.
Основная функция эпиблемы - всасывание, избирательное поглощение из почвы воды с растворенными в ней элементами минерального питания. Через эпиблему выделяется ряд веществ, например, кислот, действующих на субстрат и преобразующих его.
Цитологические особенности эпиблемы связаны с ее функциями. Это тонкостенные клетки, лишенные кутикулы, с вязкой цитоплазмой, с большим числом митохондрий (активное поглощение веществ происходит с затратой энергии).
Поглощающая поверхность эпиблемы увеличивается в 10 раз и более за счет образования корневых волосков. Корневой волосок представляет собой вырост клетки длиной 1...2 (3) мм.
При образовании корневого волоска наружная стенка клетки выпячивается, ядро перемещается в его растущий конец, где располагается в постенной цитоплазме. Здесь же находятся многочисленные диктиосомы аппарата Гольджи, продуцирующие вещества для построения клеточной стенки. Центральная вакуоль занимает большую часть клетки. Продолжительность жизни клеток эпиблемы до 15...20
Поговорим о вторичных покровных тканях…
Что представляет собой перидерма?
Перидерма (пробка или феллема) - (от греч. «пери» - вокруг и «дерма» - кожа).
Сплошная многослойная вторичная покровная ткань стеблей и корней многослойных растений.
Образуется из феллогена , который возникает из клеток основной паренхимы, лежащей под эпидермой. В процессе формирования перидермы наружу откладываются клетки феллемы, а внутрь - живые паренхимные по форме клетки - феллодермы. Пробка состоит из таблитчатых в начале живых, затем мертвых клеток, лишенных межклетников.
Их оболочка пропитана суберином. Клетки пробки воздуха - и водонепроницаемы. Она образует защитный футляр, предохраняющий живые ткани от потери воды. У пробкового дуба, бархата амурского формируется мощный слой пробки. Еѐ используют как укупорочный материал.
В перидерме с самого начала формируются чечевички - отверстия, покрытые рыхлой тканью. Через них осуществляется «проветривание» стебля, они имеют вид небольших бугорков на поверхности молодых побегов. Строение чечевичек используется в диагностики растительного сырья.
Третичная покровная ткань….
Корка – это…
В многолетних осевых органах растений развивается несколько перидерм. Постепенно они отмирают и образуют мощный покровный комплекс- корку или «ритидом». Она формируется на стволах многолетних деревьев и на корнях.
Как образуется корка?
На стволах развивается несколько перидерм, каждая последующая закладывается глубже предыдущей. Живые ткани, заключенные между слоями пробки, отмирают, и формируется покровный комплекс - корка.
Корка состоит из нескольких слоев пробки и заключенных между ними отмерших тканей.
Типы образуемых корок:
Если образование перидерм происходит не по всей окружности ствола, а отдельными полудугами, то корка формируется неправильными кусками. Такая корка называется чешуйчатой и образуется у большинства растений.
Кольцеобразная корка формируется в том случае, если каждая вновь возникающая перидерма опоясывает ствол, периодически отрезая цилиндрические участки коры (например, у винограда).
Корка не способна к растяжению, поэтому при утолщении ствола в ней появляются трещины. На дне трещин во внутренней перидерме имеются чечевички, обеспечивающие газообмен.
Также выполняет защитную роль: защищает от ожога, резких смен температуры, холода, болезни.