Алле́ли (от греч. ἀλλήλων - друг друга, взаимно) - различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологичных хромосом и определяющие альтернативные варианты развития одного и того же признака. В диплоидном организме может быть два одинаковых аллеля одного гена, в этом случае организм называется гомозиготным, или два разных, что приводит к гетерозиготному организму. Термин «аллель» предложен В. Иогансеном(1909 г.) .
Нормальные диплоидные соматические клетки содержат два аллеля одного гена (по числу гомологичных хромосом), а гаплоидные гаметы - лишь по одному аллелю каждого гена. Для признаков, подчиняющихся законам Менделя, можно рассматривать доминантные и рецессивные аллели. Если генотип особи содержит два разных аллеля (особь - гетерозигота), проявление признака зависит только от одного из них - доминантного. Рецессивный же аллель влияет нафенотип, только если находится в обеих хромосомах (особь - гомозигота). Таким образом, доминантный аллель подавляет рецессивный. В более сложных случаях наблюдаются другие типы аллельных взаимодействий (см. ниже) .
Следует, однако, отметить, что, несмотря на разнообразие взаимодействия аллелей, порой весьма сложных, все они подчиняются первому закону Менделя - закону единообразия гибридов первого поколения .
Обозначение аллелей[править | править вики-текст]
Обычно в качестве обозначения аллеля применяют сокращение названия соответствующего гена до одной или нескольких букв. В случае наличия только 2 аллелей, чтобы отличить доминантный аллель от рецессивного, первую букву в обозначении доминантного пишут заглавной.
Если же имеет место множественный аллелизм, то используется следующая система обозначения аллелей, например, в случае вышеописанной мутации ade у дрожжей. Ответственный за неё ген обозначают заглавными латинскими буквами: ADE 2 . Множественные аллели этого гена записывают так: ade 2-1 , ade 2-2 и т.д. Сам же признак, как это принято в генетике микроорганизмов, обозначают ADE .
При этом само слово «аллель» употребляется в русском языке как в женском, так и в мужском роде .
Типы аллельных взаимодействий[править | править вики-текст]
Вскоре после переоткрытия законов Менделя было установлено, что взаимодействие генов не исчерпывается одним лишь полным доминированием доминантной аллели над рецессивной. В действительности между аллелями одного гена и различными генами в генотипе существуют сложные и разнообразные взаимодействия, играющие немаловажную роль в реализацииинформации, заключённой в каждом отдельном гене .
Доминантность[править | править вики-текст]
Основная статья: Доминантность
примере цветкарододендрона (Rhododendron ).
Доминирование (доминантность) заключается в том, что один из аллелей пары (доминантный) маскирует или полностью подавляет проявление второго аллеля (рецессивного). При этом степень подавления рецессивного признака обусловливает различные варианты доминирования:
1. Полное доминирование - взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантной аллелью. Например, в экспериментах Менделя пурпурная окраска цветкаполностью доминировала над белой .
2. Неполное доминирование - доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака. Например, если в гомозиготном состоянии один аллель определяет красную окраску цветка, а другой - белую, то гетерозиготный гибрид будет иметь розовые цветки . В некоторых источниках неполное доминирование характеризуют как такой тип взаимодействия аллелей, когда признак у гибридов F 1 занимает не среднее положение, а отклоняется в сторону родителя с доминирующим признаком. Полностью же средний вариант (как, например, приведённый выше пример наследования окраски цветков) относят кпромежуточному характеру наследования , то есть отсутствию доминирования .
3. Сверхдоминирование - более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. На этом типе аллельного взаимодействия основано явление гетерозиса(превосходство над родителями по жизнеспособности, энергии роста, плодовитости, продуктивности) .
4. Кодоминирование - проявление у гибридов нового варианта признака, обусловленного взаимодействием двух разных аллелей одного гена. При этом, в отличие от неполного доминирования, оба аллеля проявляются в полной мере. Наиболее известным примером является наследование групп крови у человека . Некоторые источники также понимают именно под кодоминированием отсутствие доминантно-рецессивных отношений .
5. Доминирование, связанное с полом происходит тогда, когда одна и та же аллель у самцовпроявляется как доминантная, а у самок - как рецессивная. Например, у овцематок доминирует комолость (R), а у баранов - рогатость (R 1) .
Генные мутации.
Генные мутации
Генные (точковые) мутации затрагивают, как правило, один или несколько нуклеотидов, при этом один нуклеотид может превратиться в другой, может выпасть(делеция), продублироваться, а группа нуклеотидов может развернутся на 180 градусов. Например, широко известен ген человека, ответственный за серповидно – клеточную анемию, который может привести к летальному исходу. Соответствующий нормальный ген кодирует одну из полипептидныз цепей гемоглобина. У мутантного гена нарушен всего один нуклеотид (ГАА на ГУА). В результате в цепи гемоглобина одна аминокислота заменена на другую(вместо глутамина – валин). Казалось бы ничтожное изменение, но оно влечет за собой роковые последствия: эритроцит деформируется, приобретая серповидно – клеточную форму, и уже не способен транспортировать кислород, что и приводит к гибели организма. Генные мутации приводят к изменению аминокислотной последовательности белка. Наиболее вероятное мутация генов происходит при спаривание тесно связанных организмов, которые унаследовали мутантный ген у общего предка. По этой причине вероятность возникновения мутации повышается у детей, чьи родители являются родственниками. Генные мутации приводят к таким заболеваниям, как амавротическая идиотия, альбинизм, дальтонизм и др.
Интересно, что значимость нуклеотидных мутаций внутри кодона неравнозначна: замена первого и второго нуклеотида всегда приводит к изменению аминокислоты, третий же обычно не приводит к замене белка. К примеру, "Молчащая мутация"- изменение нуклеотидной последовательности, которая приводит к образованию схожего кодона, в результате аминокилотная последовательность белка не меняется.
Хромосомные мутации
Хромосомные мутации приводят к изменению числа, размеров и организации хромосом, поэтому их иногда называют хромосомными перестройками. Хромосомные перестройки делятся на внутри- и межхромосомные. К внутрехромосмным относятся:
· Дубликация – один из участков хромосомы представлен более одного раза.
· Делеция – утрачивается внутренний участок хромосомы.
· Инверсия –повороты участка хромосомы на 180 градусов.
Межхромосомные перестройки (их еще называют транслокации) делятся на:
· Реципрокные – обмен участками негомологичных хромосом.
· Нереципрокные – изменение положения участка хромосомы.
· Дицентрические – слияние фрагментов негомологичных хромосом.
· Центрические – слияние центромер негомологичных хромосом.
Хромосомные мутации проявляются у 1% новорожденных. Однако интересно, исследования показали, что нестабильность соматических клеток здоровых доноров не исключение, а норма. В связи с этим была высказана гипотеза о том, что нестабильность соматических клеток следует рассматривать не только как патологическое состояние, но и как адаптивную реакцию организма на измененные условия внутренней среды. Хромосомные мутации могут обладать фенотипическими явлениями. Наиболее распостраненный пример - синдром "Кошачьего крика" (плачь ребенка напоминает мяукание кошки). Обычно носители такой делеции погибают в младенчестве. Хромосомные мутации часто приводят к паталогическим нарушениям в организме, но в то же время хромосомные перестройки сыграли одну из ведущих ролей в эволюции. Так, у человека 23 пары хромосом, а у обезьяны - 24. Таким образом различие составляет всего одна хромосома. Ученые предполагают, что в процессе эволюции произошла хотя бы одна перестройка. Подтверждением этого может служить и тот факт, что 17 хромосома человека отличается от такой же хромосомы шимпанзе лишь одной перецентрической инверсией. Такие рассуждения во многом подтверждают теорию Дарвина.
Геномные мутации
Главная отличительная черта геномных мутаций связана с нарушением числа хромосом в кариотипе. Эти мутации так же подразделяются на два вида: полиплоидные анеуплоидные.
Полиплоидные мутации ведут к изменению хромосом в кариотипе, которое кратно гаплоидному набору хромосом. Этот синдром впервые был лишь обнаружен в 60-ых годах. Вообще полиплодия характерна в основном для человека, а среди животных встречается крайне редко. При полиплоидии число хромосом в клетке насчитывается по 69 (триплодие) , а иногда и по 92 (тетраплодие) хромосомы. Такое изменение ведет практически к 100 % смерти зародыша. Триплодие имеет не только многочисленные пороки, но и приводит к потере жизнеспособности. Тетраплодие встречается еще реже, но так же зачастую приводит к летальному исходу.
Анеуплоидные же мутации приводят к изменению числа хромосом в кариотипе, некратное гаплоидному набору. В результате такой мутации возникают осыби с аномальным чилом хромосом. Как и триплодия, анеуплодия часто приводит к смерти еще на ранних этапах развития зародыша. Причиной же таких последствий является утрата целой группы сцепления генов в кариотипе.
В цело же, механизм возникновения геномных мутаций связан с патологией нарушения нормального расхождения хромосом в мейозе, в результате чего образуются аномальные гаметы, что и ведет к мутации. Изменения в организме связаны с присутствием генетически разнородных клеток. Такой процесс называется мозаицизм.
Геномные мутации одни из самых страшных. Они ведут к таким заболеваниям, как синдром Дауна (трисомия, возникает с частотой 1 больной на 600 новорожденных), синдром Клайнфельтера и др.
Классификация мутаций по уровню организации наследственной информации:
- Генные (точковые ) – невидны в световой микроскоп. Причиной может быть выпадение нуклеотида, повтор одного и того же нуклеотида, замена одного нуклеотида на другой. Это всё приводит к изменению наследования нуклеотидов в молекулах ДНК. А это в свою очередь приводит к изменению последовательности аминокислот в молекуле белка.
- Хромосомные(аберрации ) – изменение макроструктуры хромосом. видны в световой микроскоп. Различают несколько видов хромосомных мутаций.
Межхромосомные (транслокации) – перенос целой хромосомы или её части и присоединение этого участка к другой негомологичной хромосоме.
Внутрихромосомные (инверсии) – выпадение участка хромосомы. Разворот этого участка хромосомы на 180 градусов и вновь встраивание в эту же хромосому.
Делеция (выпадение) – потеря срединного участка хромосомы.
Дефишенси - потеря концевого участка хромосомы.
Дупликация – повтор одного и того же участка хромосомы.
- Геномные – обусловлены изменением числа хромосом. Сами хромосомы при этом не изменяются.
Полиплоидия – увеличение числа хромосом на набор кратный гаплоидному (3n, 4n, 5n).
< Аутоплоидия – организмы, в которых умножаются хромосомы одного генома.
< Аллоплоидия – организмы, в которых умножаются хромосомы разных геномов.
Гетероплоидия – происходит изменение числа хромосом на набор не кратный гаплоидному (2n+1 - трисомия, 2n-1 - моносомия, 2n-2 - нулисомия).
Гаплоидия – в соматических клетках имеется только гаплоидный набор хромосом.
Цитоплазматические – возникают в ДНК органелл цитоплазмы.
Классификация мутаций по типу клеток, в которых возникают мутации (по локализации):
- Генеративные – возникают половых клетках и наследуются при половом размножении.
- Соматические – возникают в соматических клетках, не наследуются, но могут наследоваться у растений при вегетативном размножении.
Классификация мутаций по причине их вызывающей:
- Спонтанные – причина неизвестна.
- Индуцированные – причина известна.
Мутагены – это факторы, вызывающие мутации.
Вы когда-нибудь задумывались, почему у вас есть такой особый цвет глаз или тип волос? Все это связано с передачей генов. Как обнаружил Грегор Мендель, черты наследуются при помощи передачи генов от родителей к их потомкам. представляют собой участки ДНК, расположенные на наших . Они передаются от одного поколения к другому через . Ген для конкретного признака может существовать в более чем одной форме или аллеле. Для каждой характеристики или признака обычно наследуют два аллеля. Парные аллели могут быть гомозиготными (с идентичными аллелями) или гетерозиготными (с разными аллелями) для данного признака.
Когда пары аллелей одинаковы, генотип этого признака идентичен, а или характеристика, которая наблюдается, определяется гомозиготными аллелями. Когда парные аллели для признака различные или гетерозиготные, возможно несколько вариантов. Гетерозиготные аллели, которые обычно наблюдаются в клетках животных, включают полное доминирование, неполное доминирование и кодоминирование.
Полное доминирование
При этом виде доминирования один аллель является доминирующим, а другой рецессивным. Доминантный аллель полностью маскирует рецессивный. Фенотип определяется доминирующим аллелем. Например, гены для формы семени в растениях гороха существуют в двух формах: гладкая (R) и морщинистая (r). В растениях гороха, которые являются гетерозиготными по форме семени, гладкий горох является доминирующим по отношению морщинистому семени, а генотип является (Rr).
Неполное доминирование

При неполном доминировании один аллель для конкретного признака не является полностью доминирующим над другим аллелем. Это приводит к третьему фенотипу, в котором наблюдаемые характеристики представляют собой смесь доминирующих и рецессивных фенотипов. Пример неполного доминирования проявляется в наследовании типа волос. Кудрявый тип волос (CC) является доминирующим для прямого типа волос (сс). Человек, гетерозиготный по этому признаку, будет иметь волнистые волосы (Cc).
Доминирующая фигурная характеристика не полностью выражена по прямой характеристике, создавая промежуточную характеристику волнистых волос. При неполном доминировании одна характеристика может быть несколько более заметной, чем другая для данного признака. Например, у человека с волнистыми волосами может быть больше или меньше волн, чем у другого с волнистыми волосами. Это указывает на то, что аллель для одного фенотипа выражается немного больше, чем аллель для другого фенотипа.
Кодоминирование

Нормальная и серповидная формы эритроцита
При совместном доминировании ни один из аллелей не является доминирующим, но оба аллеля для конкретного признака полностью выражены. Это приводит к третьему фенотипу, в котором наблюдается более одного фенотипа. Пример кодоминирования наблюдается у индивидов с чертой серповидной клетки.
Наличие серповидной клетки связано с развитием эритроцитов с аномальной формой. Нормальные эритроциты имеют двояковогнутую, дискообразную форму и содержат огромное количество белка, называемого гемоглобином. Гемоглобин помогает красным клеткам связываться и переносить кислород в клетки и ткани организма. Серповидная клетка является результатом мутации гена гемоглобина. Этот гемоглобин считается ненормальным и заставляет клетки крови принимать серповидную форму.
Серповидные клетки часто застревают в кровеносных сосудах и блокируют нормальный кровоток. Те, которые несут черту серповидной клетки, гетерозиготные для гена гемоглобина и наследуют один нормальный ген гемоглобина и один серповидный ген гемоглобина. У них нет болезни, потому что аллель гемоглобина серпа и нормальный гемоглобиновый аллель являются кодоминантными по отношению к форме клеток. Это означает, что у носителей серповидных клеток образуются как нормальные эритроциты, так и серповидные.
Неполное доминирование и кодоминирование

Розовый цвет тюльпанов представляет собой смесь выражения обоих аллелей (красного и белого), в результате чего образуется промежуточный фенотип (розовый). Это неполное доминирование. В красно-белом тюльпане оба аллеля полностью выражены. Это свидетельствует о кодоминировании.
Люди часто путают неполное доминирование и кодоминирование. Хотя они и являются примерами наследования, но отличаются выражением генов. Ниже перечислены некоторые различия между ними:
Экспресивность аллеля
- Неполное доминирование: один аллель для определенного признака не полностью выражен над его парным аллелем. Например, цвет тюльпана, аллель для красного цвета (R) не полностью маскирует аллель для белого цвета (r).
- Кодоминирование: оба аллеля для конкретного признака полностью выражены. Аллель для красного цвета (R) и аллель для белого цвета (r) выражены и видны в гибриде.
Зависимость аллелей
- Неполное доминирование: влияние одного аллеля зависит от его парного аллеля для данного признака.
- Кодоминирование: влияние одного аллеля не зависит от его парного аллеля для данного признака.
Фенотип
- Неполное доминирование: Гибридный фенотип представляет собой смесь обеих аллелей, что приводит к третьему промежуточному фенотипу. Пример: Красный цветок (RR) X Белый цветок (rr) = Розовый цветок (Rr)
- Кодоминирование: гибридный фенотип представляет собой комбинацию выраженных аллелей, что приводит к третьему фенотипу, который включает оба фенотипа. Пример: Красный цветок (RR) X Белый цветок (rr) = Красно-белый цветок (Rr)
Наблюдаемые характеристики
- Неполное доминирование: фенотип может быть выражен в разной степени в гибриде. (Пример: розовый цветок может иметь более легкую или более темную окраску в зависимости от количественной выраженности одного аллеля по сравнению с другим).
- Кодоминирование: оба фенотипа полностью выражены в гибридном генотипе.
Краткий вывод
При неполном доминировании один аллель для конкретного признака не является полностью доминирующим над другим аллелем. Это приводит к третьему фенотипу, в котором наблюдаемые характеристики представляют собой смесь доминирующих и рецессивных фенотипов.
При кодоминировании ни один аллель не является доминирующим, но оба аллеля для конкретного признака полностью выражены. Это приводит к третьему фенотипу, в котором наблюдается более одного фенотипа.
Чтобы понять, что же такое кодоминирование в генетике, разберем возможные типы взаимодействия Согласно гипотезе чистоты гамет, предложенной Грегором Менделем, при образовании гаметы в нее попадает только один из двух аллельных генов каждого родительского организма, отвечающий за этот признак. Так в гамете образуется нормальный диплоидный набор аллельных генов. Далее во взаимодействии может проявиться полное доминирование, когда подавит рецессивный, неполное доминирование и кодоминирование.
Неполное доминирование
В этом случае доминантный аллель не полностью подавляет рецессивный, в результате получается новый, промежуточный признак. Известным примером неполного доминирования является окраска цветков некоторых цветов, например космеи. Допустим, есть гомозиготный красный цветок с генотипом (АА) (чистая линия) и белый цветок (аа), тоже чистая линия. При их скрещивании появляются цветы с розовой окраской - пример кодоминирования. Их генотип имеет вид Аа, но и доминантный, и рецессивный аллель проявляются. При скрещивании получился промежуточный - розовая окраска.
Кодоминирование
Другой тип экспрессии генов - кодоминирование. Это явление похоже на неполное доминирование, но все же имеет одно существенное отличие. Кодоминирование - взаимодействие генов, при котором противоположные признаки проявляются одновременно, но не смешиваются и не производят промежуточный признак. 
При скрещивании белого цветка петунии с красным может получиться красный, розовый, белый или двухцветный. Цветок с красными и белыми полосами появляется в результате такого процесса, как кодоминирование. Это самый распространенный пример такого взаимодействия.

Кодоминирование характерно и для других растений.
Взаимодействие неаллельных генов
Стоит сказать, что только к аллельным генам применимы такие понятия, как и кодоминирование.  Примеры и многочисленные эксперименты подтверждают, что в случае неаллельных генов называют другие типы взаимодействия - кооперация, эпистаз, комплементарность, полимерия. Примером именно полимерии, а не неполного доминирования, является наследование цвета кожи человека.
Примеры и многочисленные эксперименты подтверждают, что в случае неаллельных генов называют другие типы взаимодействия - кооперация, эпистаз, комплементарность, полимерия. Примером именно полимерии, а не неполного доминирования, является наследование цвета кожи человека.
Кодоминирование у человека
Другой простой, но яркий пример кодоминирования - наследование групп крови. Как известно, существует четыре группы крови. Первая группа О(І) проявляется при наличии в генотипе двух гомозиготных рецессивных генов О. вторая группа А(ІІ) может проявиться и при генотипе АО или АА. В фенотипе при этом будет проявляться только доминантный ген А, который полностью подавит рецессивный ген. Похожая ситуация будет и для третьей группы крови В(ІІІ), которая формируется при генотипе ВВ или ВО. В подавит рецессивный ген О и проявится как результат полного доминирования. Но что будет при скрещивании гомозигот с генотипами АА и ВВ? И ген А, и ген В доминантны, значит ни один из них не может полностью подавить другой и проявится самостоятельно. В этом случае с вероятностью 100% получится четвертая группа крови - АВ, имеет место кодоминирование. Это же происходит при скрещивании гетерозигот АО и ВО, когда возможен любой результат:
F1: АО(II), АВ(IV), ВО(III), ОО(I).
Именно поэтому группа крови ребенка может не совпадать с группой крови родителей. Из примера видно, что кодоминирование проявляется не только в окраске растений.
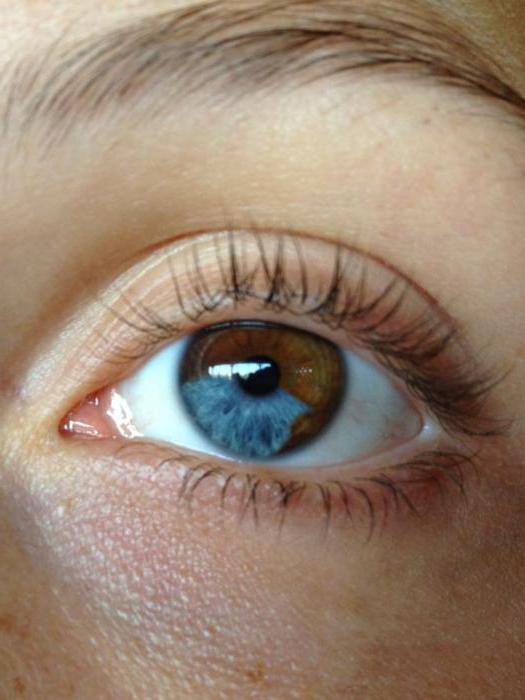
Кодоминорование и мутации
Стоит оговориться, что проявление обоих признаков - это не всегда кодоминирование. Это доказывает редкая генетическая особенность, свойственная людям и некоторым животным - гетерохромия (несовпадение окраски радужной оболочки глаз). Гетерохромия бывает полная, например, когда один глаз карий, а второй голубой, или частичной, например когда на зеленой оболочке есть серый сегмент. Гетерохромия, несмотря на кажущуюся аналогию с окраской цветов, пример не кодоминирования, а Нарушение пигментации кожи - также не кодоминирование, о чем говорит генетика. Кодоминирование в этом случае путают с заболеваниями.
Кодоминирование и первый закон Менделя
Явления кодоминирования и неполного доминирования, на первый взгляд, говорят о том, что первый о единообразии гибридов не выполняется. Грегор Мендель в своих экспериментах имел дело с горохом, для которого не свойственно ни кодоминирование, ни частичное доминирование, а только полное доминирование. В тех случаях, если смешанный признак или их одновременное проявление невозможно, его формулировка была абсолютно правильной. Спустя почти столетие, когда были исследованы и кодоминирование, и неполное доминирование, в первый закон была внесена поправка, гласившая, что при скрещивании гомозиготных гибридов первого поколения с противоположными признаками во втором поколении появляются гибриды, по этому признаку идентичные. Проявляется доминантный признак в случае полного доминирования или смешанный признак - в случае неполного доминирования.
Можно воспользоваться примером с наследованием группы крови, чтобы наглядно продемонстрировать правильность дополненного первого закона Менделя:
F1: АВ, АВ, АВ, АВ.
Результатом скрещивания двух чистых линий будет гетерозиготная особь, в фенотипе которой проявляется смешанный признак, так как имеет место кодоминирование. Это соответствует внесенной поправке.
Генетика
1. Предмет и цель генетики. Понятие о наследственности, наследовании и изменчивости.
2. Основные методы исследования применяемые в генетике.
3. Синтез белка в клетке. Передача наследственной информации в системе ДНК, РНК, белок.
4. Ядро. Диплоидный и гаплоидный набор хромосом.
5. Расщепление при моно- и дигибридном скрещивании и соотношение генотипов и фенотипов для этих скрещиваний.
6. Митотический цикл в период интерфазы.
7. Закономерности наследования признаков установленные Менделем.
8. Понятие о кариотипе и генотипе. Аутосомы и половые хромосомы.
9. Летальные гены и их действия.
10. Дигибридное скрещивание, особенности гибридов 1-го и 2-го поколения.
11. Виды доминирования: полное, как объяснить явления доминирования.
12. Строение и роль хромосом в передаче наследственной информации.
13. Понятие о сверх- и кодоминирповании.
14. Мейоз, его генетическая сущность.
15. Гибридологический метод изучения наследственности разработанный Менделем. Его особенности.
16. Моногибридное скрещивание и его схема. Изучение наследования признаков в поколении.
17. Строение генов и их влияние на развитие признаков.
33. Типы хромосомного определения пола у животных: сингамное, эпигамное, прогамное.
39. Сцепленное наследование. Используя конкретный пример, составить схему скрещивания и охарактеризуйте его.
40. Понятие об изменчивости. Виды изменчивости: модификационная изменчи
вость.
44. Спонтанные и индуцированные мутации. Мутагенные факторы. Частота и причины мутаций.
1. Предмет и цель генетики. Понятие о наследственности, наследовании и изменчивости.
Термин был предложен в 1906 году английским биологом У. Бэтсоном и в переводе генетика означает «рождение или происхождение». Генетика - это наука о наследственности и изменчивости всех живых организмов. Цель состоит в изучении закономерностей наследованностей и изменчивостей и разработки методов практического использования этих закономерностей. В 1900 году вторично открыты законы Менделя тремя учёными: в Голландии Дэ Фризом, в Германии - Корренс, в Австрии - Чермак. Соновоположником генетики является Чешский учёный учёный Мендель, но большой вклад в изучение современной генетики внесли: американский генетик Томас Морган и его школа. Которые разработали хромосомную теорию наследственности в 1910 году. В 20 - 30 -х годах вклад в генетику внесли работу советских учёных: Вавилов, Кольцов, Четвериков, Серебровский. Середины 30 -х годолв после 48 года в генетике преобладали антинаучные взгляды Лысенка. В настоящее время генетика развивается интенсивно и в нашей стране ведут в области генетики 200 институтов. Наследственность - это св-ва организмов передавать свои признаки потомству. В генетике понятие признак употребляют для обозначения характерных особенностей, которые отличают одну особь от другой. Наследование - это процесс, способ передачи генетической информации от ролителей к потомкам. Изменчивость - это св-во живых организмов изменяться. Изменчивость выражается у различных потомков и родителей. Наследуемость - это доля генетической изменчивости в общей фенотипической изменчивости признака для популяции или группы организмов.
2. Основные методы исследования применяемые в генетике.
Генетика изучает наследственность и изменчивость качественных и количественных признаков. Качественные признаки оцениваются визуально (масть, упитанность). Количественные призанки - это те признаки, которые измеряются и выражаются числовыми значениями (удой, масса). Генетическое многообразие проявления признаков обуславливают и множество методов, современное изучение наследственности и изменчивости осуществляется путём генетиечского анализа - это совокупность методов исследования наследственных св-в организма. К основным методам генетического анализа относятся : Гибридологический метод сущность его в том, чтобы получить потомков от родительских особей отличающихся друг от друга по одним или нескольким призанкам. Статистический или биометрический . Применяется при математической обработке результатов скрещиваний при изучении групповых св-в биологических объектов. Этот метод порзволяет определить соответствуют ли полученные экспериментальные данные теоритическиожидаемым результам. Этот метод применяется при изучении связей между признаками. Генеалогический метод . Этот метод требует составления родословных, т.е. сведений о предков и потомков разных поколений. Биохимический метод служит для изучения передачи наследственной информации, как работают гены. Цитологический служит для изучения строения, функционирования хромосом, т.е. позволяет выявить наследственные болезни. Биотехнологический . Этот метод использования живых организмов и бологических процессов в производстве для получения необходимых человеку веществ. Этот метод используется для получения витаминов, ферментов, гормонов, трансплантация эмбрионов, клонирование.
3. Синтез белка в клетке. Передача наследственной информации в системе ДНК, РНК, белок.
Процесс синтеза белка в клетке называется биосинтезом . Он осуществляется под контролем ДНК, котрое реализует через синтез белка заложенную в ней генетическую информацию: ДНК à[и-РНК, р-РНК, т-РНК] àбелки àпризнак. Основные этапы синтеза : 1)Транскрипция. 2)Трансляция. Транскрипция означает копирование. При транскрипции на определённом участке молекулы ДНК синтезируется и-РНК из нуклеотидов, порядок которых точно скопирован с этого участка молекулы ДНК. Транскрипция только в ядре происходит. и-РНК отделяется от ядра и из ядра перемещается в цитоплазму и из цитоплазмы к рибосоме. Трансляция осуществляется на рибосомах. В неё и-РНК определяет порядок соединения аминокислот при синтезе белковой молекулы. К рибосоме доставляются аминокислоты т-РНК, а рибосомы участвуют в сборке белковой молекулы. За 1 секунду удленяется полипептидная цепь на 7 аминокислот. В среднем синтез 1 молекулы белка происходит в течении 3-4 секунд. В результате половина белком нашего тела обновляется за 80 дней. За всю свою жизнь человек обновляет весь свой белок около 200 раз. Условия для биосинтеза : Наличие энергии. энергия накапливается в митохондриях в виде АТФ. Соответствующие ферменты. Наличие информации о структуре белка. Информация в ДНК. Наличие аминокислот. Наличие рибосом, как фабрик белка с помощью р-РНК.
4. Ядро. Диплоидный и гаплоидный набор хромосом.
Ядро - это важный компонент клеток эукариот. Большинство клеток имеют 1-но ядро, но встречаются многоядерные клетки. У одноклеточных (скелетные мышцы у позвоночных) некоторые клетки утрачивают ядра (эритроциты млекопитающих). Ядра имеют шаровидную или овальную форму. Ядро интерфазное, т.е. сотоит из неделящихся клеток. Ядро состоит из : Ядерной мембраны. Кариоплазмы, т.е. ядерного сока. Ядрышка. Хроматина. Ядерная мембрана образована 2-мя мембранами: наружной и внутренней и содержит многочисленные поры, через которые между ядром и цитоплазмов происходит обмен различных в-в. Кариоплазма представляет собой желеобразный р-р, в котором находится нуклеотиды, белки и другие в-ва. Ядрышки одно или несколько представляют собой тельца округлой формы. Функция ядрышка заключается в синтезе р-РНК. Ядрышки непостоянные ядерной структуры. Они исчезают в начале деления и появляются в конце деления и они Хроматин хроматиды центромером плечами хромосом
Строение хромосомы хромонема » или хромотиновая нить гистона Например
гомологичными (2 N ) (N ) .
5. Расщепление при моно- и дигибридном скрещивании и соотношение генотипов и фенотипов для этих скрещиваний.
Закономерности: 1) 1-ый закон Менделя 2) Закон расщепления или 2-й закон Менделя Ф=3:1 , расщепление по Г=1:2:1 . 3) 3-й закон Менделя Ф=9:3:3:1 Г=1:2:1:2:4:1:2:2:1=16 гибридов.
6. Митотический цикл в период интерфазы.
МИТОЗ (гр. Нить) - не прямое деление. Происходит путём сложного деления ядра и цитоплазмы. Митозом делятся соматические клетки, происходит дробление зиготы, образование зародышевых клеток. Митоз составляет часть митотического цикла клетки. Митотический цикл - это существование клетки от одного деления до другого деления. Он состоит из интерфазы и митоза. ДЕЛЕНИЕ ИНТЕРФАЗЫ : G 1 - постмитотический или пресинтетический, S - период синтеза, G 2 - постсинтетический или премитотический. G S - период .Происходит синтез ДНК, т,е. Удвоение молекулы ДНК. Происходит построение 2-й хроматиды, в которую переходит вновь синтезированная молекула ДНК, образуется двухроматидная хромосома. В этот же пероид синтезируются белки - “гистоны”. G 2 - период, в котором :1)Вновь происходит синтез и накопление энергии в виде АТФ. 2)Клетка синтезирует специфические белки, из которых образуются нити веретена деления. МИТОЗ - период деления. Это сложный непрерывный процесс в котором образуются условно 4-и стадии: Профаза, Метафаза, Анафаза, Телофаза. У большинства клеток весь митоз происходит в течении 1-2 часов.
7. Закономерности наследования признаков установленные Менделем.
С помощью гибридологического метода на основании экспериментального анализа гибридов Мендель установил закономерности наследования признаков в 1865 г. Закономерности: 1) Закон единообразия гибридов 1-го поколения или 1-ый закон Менделя . Он был установлен на основе моногибридного скрещивания «При моногибридном скрещивании все потомки 1-го поколения единообразны и наследуют доминантный признак одного из родителей независимо у какого родителя (отца или матери) был этот признак». 2) Закон расщепления или 2-й закон Менделя : «При скрещивании гибридов 1-го поколения между собой, во 2-ом поколении происходит расщепление в определённом количественном соотношении (Доминантных - 3, рецессивных - 1 признак). Во 2-ом поколении образуется 2-е фенотипические группы расщепления по Ф=3:1 , расщепление по Г=1:2:1 . 3) Закон независимого наследования или 3-й закон Менделя . На основе дигибридного скрещивания. «При дигибридном скрещивании каждая пара признаков наследуется независимо от других. Во 2-ом поколении образуется 4-е фенотипические группы. Расщепление по Ф=9:3:3:1 и во 2-ом поколении образуется 9-ть разных генотипов Г=1:2:1:2:4:1:2:2:1=16 гибридов.
8. Понятие о кариотипе и генотипе. Аутосомы и половые хромосомы.
Кариотипом называется совокупность хромосомного набора (это размер, число и форма) соматических клеток данного вида. Кариотип строго специфичен для каждого вида. Хромосомы в составе кариотипа делятся на аутомомы и гетеросомы (половые хромосомы). Аутосомы - одинаковые хромосомы в кариотипе самцов и самок. Дрозофила - 8А. Гетеросомы
положения центромера. Геном - это совокупность хромосом и генов присущих половым клеткам, т.е. гаплоидный набор хромосом.
9. Летальные гены и их действия.
Плейотропия Примеров много: у человека один и тот же ген определяет цвет волос (рыжий), веснушки. Практически каждый ген контролирует определённый этап метаболизма. Многоэталонность метаболизма приводит к тому, что нарушение метаболизм на одном этапе неизбежно сказывается на признаках. Часто плейотропные гены вызывают видимые внешние морфологические изменения, но в внутренние строение внутренних органов; и в итоге сказываются на жизнеспособность или привести организм к гибели. Гены приводящие организм к гибели на определённой стадии онтогенеза смерти называют летальными генами. Летальные гены летальные гены обладают двойным действием: с одной стороны вызывают развитие полезного для человека хозяйственно-полезного признака (окраска животных). С другой стороны со стороны плейотропии снижает жизнеспособность и даже приводит к гибели. Среди овец каракульской породы встречаются животные с красивой серой серебристой окраской (Аа ûАа). Ген - доминантный
Примером плейотропного действия гена у человека служит рецессивная мутация в гене, которая контролирует синтез в полипептидной цепи в молекуле гемоглобина. Мутация вызывает замену одной аминокислоты и это влечёт в изменении морфологии эритроцитов (серповидной формы), к нарушению сердечно-сосудистой, нервной систем и т.д. Гомозиготность приводит по этой мутации к гибели. Часто летальные гены являются рецессивными, поэтому гетерозиготы нормальны по фенотипу, а у рецессивных гомозигот они проявляются. Летальные гены встречаются у животных всех видов в аутосомах и половых хромосомах. К настоящему времени выявлено у КРС 46 летальных генов; кур - 45; Лошадь - 10; Свинья - 18; Овца - 15. Модифицирующее действие генов: признак под контролем: гена модификатора; основного. Пример: Модифицирующего действия белая пятнистость шерсти у УКРС. Ген основного действия
10. Дигибридное скрещивание, особенности гибридов 1-го и 2-го поколения.
3) Закон независимого наследования или 3-й закон Менделя . На основе дигибридного скрещивания. «При дигибридном скрещивании каждая пара признаков наследуется независимо от других. Во 2-ом поколении образуется 4-е фенотипические группы. Расщепление по Ф=9:3:3:1 и во 2-ом поколении образуется 9-ть разных генотипов Г=1:2:1:2:4:1:2:2:1=16 гибридов.
11. Виды доминирования: полное, как объяснить явления доминирования.
Развитие любых признаков у организмов является результатом сложного взаимодействия между генами, точнее между белками, которые синтезируются под контролем генов. Аа, аа, АА Взаимодействие генов внутри аллельной пары: Полное, неполное, сверхдоминирование, кодоминирование. АаВв Взаимодействие генов между разными аллельными генами: Комплиментарное, эпистаз, полимерия. Полное доминирование - это преобладание одних признаков над другими. Полное доминирование проявляется в тех случаях, когда одна аллель гена (доминантная) полностью подавляет другой аллель. При полном доминировании гибрида 1-го поколения по фенотипу единообразны между собой и с родителем имеющий доминантный признак. При скрещивании гибридов 1-го поколения между собой во 2-ом поколении произойдёт расщепление по фенотипу 3:1 . Явление доминирования объясняется взаимодействием внутри одной пары аллелей, т.е. это аллельное взаимодействие. Биохимики показали, что у гомозигот оба аллеля контролируют синтез одного и того же белка-фермента. У гетерозигот в случае полного доминирования возможны 2-а случая объяснения: 1)Белок-фермент , который синтезируется под контролем доминантного аллеля полностью связывает белок полученный под контролем рецессивного аллеля А-чёрн. а-красн. F1 - Аа - чёрн. F2 АА; Аа; Аа; аа. Ф=3:1 Г=1:2:1. 2)белок , который синтезируется под контролем рецессивного гена Аа не активный и фенотипически не проявляется.
12. Строение и роль хромосом в передаче наследственной информации.
Ядрышко являются производными хромосом. Хроматин представляет собой плотные структуры в виде гранул, зёрен и образованы молекулами ДНК. Хроматин представляет собой дисперсное состояние хромосом в интерфазе. В процессе деления клеток происходит спирализации молекул ДНК их хроматиновые структуры образуют хромосомы. Термин «хромосома» был предложен немецким учёным Вальдейер в 1888 году. «Chromo» - красный, «Soma» - тело. Хромосомы видны только при делении, хотя они являются постоянными клеточной структуры. Хромосомы - это специализированные клеточные органоиды с помощью которых осуществляется хранение и передача в поколениях генетической информации клетки. Хромосомы лучше всего видны в стадии метафазы митоза. Каждая метафазная хромосома состоит из 2-х продольных копий, которые называются хроматиды . Хроматиды соединяются между собой центромером . Центромера делит хромосому на 2-е части, которые называются плечами хромосом . В зависимости от расположения центромер хромосомы подразделяются: Метацентрические. Субметацентрические. Акроцентрические. Спутничные.
Строение хромосомы : в доль каждой хроматиды идёт двойная спирализованная нить («хромонема » или хромотиновая нить ). Биохимически установили, что хромонема состоит из: молекул особого белка «гистона » и ДНК. Хромосомы способны к репликации, но не способны к регенерации. Каждый вид растений, животных имеет определённое и постоянное число хромосом. Число хромосом не зависит от уровня организации. У всех особей, которые относятся к одному виду число хромосом в ядрах всех клеток постоянно. Например : у молярийного плазмоида - 2-е хромосомы. У радиолярии - 800. У Дрозофилы - 8. У человека - 46. У Свиньи - 38. У Кролика - 44. У Овцы - 54. У КРС - 60. У Лошади - 64. У Собаки - 78. У Картофеля - 48. У Ячменя - 14.
Хромосомы, которые относятся к одной паре называются гомологичными . Гомологичые хромосомы имеют одинаковые размеры, одинаковую форму, состав и порядок расположения генов. Гомологичные хромосомы имеют разное происхождение. Одна получена от отца, другая от матери. Хромосомы не относящиеся к одной паре называются не гемологияными. Они всегда имеют отличия. В соматических клетках все хромосомы парные называется диспоидными (2 N ) . В гаметах набор хромосом половинный, клетки с одинарным набором хромосом называются гаплоидными (N ) .
13. Понятие о сверх- и кодоминирповании.
Сверхдоминирование проявляется иногда в тех случаях, когда гетерозигота первого поколения имеет более сильное проявление признаков, чем в гомозиготном состоянии родителей Р ♀АА х ♂аа F1 Аа. Сверхдоминирование проявляется у гибридов при скрещивании осла и кобылы (мул). Он превосходит родительские формы по долголетию и выносливости. Лошак - наоборот. Они бесплодны. Примером сверхдоминирования является гибрид осетровых - бестер (белуга и стерлядь) - скорость роста. Такое явление когда гибрид превосходит своих родителей по показателям называется гетерозис , который характеризуется повышенной способностью помесей. При объяснении причин сверхдоминирования возможны 2-а случая : 1) У гетерозигот аллели контролируют каждый свой белок, которые взаимодополняют друг друга, т.е. белки которые синтезируются под доминантными и рецессивными аллеля объясняются. 2)«Эффект дозы» , т.е. рецессивная гомозигота производит слишком мало белка, а доминантная гомозигота слишком много белка. А у гетерозигот доминантный находится в одной дозе и рецессивный - в одной дозе - это благоприятствует развитию признака. Кодоминирование или совместное . Проявляется в тех случаях, когда в 1-ом поколении имеют признаки обоих родителей. При кодоминировании оба аллеля проявляются полностью и нельзя сказать который доминантный, а какой рецессивный они равнозначны в своём проявлении. Биохимики показали, что в случае кодоминирования различные аллели контролируют синтез разных белков и эти белки не влияют друг на друга на проявление признака. По типу кодоминирования наследуются: группы крови у человека и животных; наследуются гены определяющие различие в строении белков гемоглобина, белков молока, белков яиц и белков волокон мышечных тканей. Если один из родителей имеет 2-ю группу крови с белком А, а второй 3-ю с белком В, то потомство будет иметь 4-ю группу: Iгр (0) - I 00 . Iiгр (А) - I АА или I АО. IIIгр (В) - I ВВ или I ВО. Ivгр (АВ) - I АВ.
14. Мейоз, его генетическая сущность.
Мейоз - наследственно-неравное деление. Мейоз характерен для образования гамет. Особый способ деления, состоит из 2-х последовательных клеточных делений, которые обозначаются: Мейоз I (профазаI, метафазаI, анафазаI, телофазаI), и Мейоз II (профазаII, метафазаII, анафазаII, телофазаII). В исходной ролительской клетки удвоение хромосом (репликация) происходит только 1 раз, а клетка делится дважды. В результате мейоза из 1-й диплоидной клетки образуется 4-е гаплоидных. Схема основных этапов мейоза : Мейоз I : в интерыазе перед 1-ым делением мейоза
нити веретина деления. Метазафа I : в метафазе биваленты выстраиваются по экватору. Анафаза I : в анафазе центромеры хромосом не делятся. К полюсам отходят целые хромосомы. Телофаза I : образуется ядерная оболочка и время длительности фазы ограничено. Синтез ДНК в промежутках между МейозомIи IIне происходит. Мейоз II : происходит также как и митоз. ПрофазаIIтакаяже как профаза митоза (исчезновение ядерной оболочки, ядрышка, уплотнение хромосом, формирование нитей веретина деления). МетафазаIIхромосомы состоящие из 2-х хроматид прикрепляются центромерами к нитям веретина и располагаются по экватору. АнафазаIIкаждая центромера делится также как и в митозе и хроматиды расходятся в противоположные стороны. ТелофазаIIзавершается образование ядерной мембраны вокруг каждого из гаплоидных ядер. Генетическая схема мейоза :
Сущность мейоза состоит в том, что происходят 2-а быстроследующих друг за другом деления клетки, причём: хромосомы реплицируются только 1 раз. Первое деление мейоза называется редукционным , т.е. при котором происходит уменьшение числа хромосом вдвое. В профазеIнаблюдается 2-а важных в генетическом отношении события: 1) Коньюгация гомологичных хромосом. 2) Кроссинговер. Кроссинговер обеспечивает образование новых комбинаций ♀ и ♂ генов в хромосомах будующих гамет. Второе деление мейоза (эквационное или уравнительное) схожа в митозом. Здесь центромера каждой хромосомы, сотоящей из 2-х хроматид делится и дочерние хромосомы в равном но уменьшенном вдвое количестве расходятся в новые клетки. Генетическое значение мейоза : 1)Уменьшение числа хромосом. 2)Возникновение новых типов хромосом.
15. Гибридологический метод изучения наследственности разработанный Менделем. Его особенности.
Гибридологическим методом называется метод скрещивания родительских организмов размножающихся по отдельным признакам и изучения проявления этих признаков у потомства в ряду поколения. Особенности гибридологического метода: Изучались наследования отдельных конкретных (альтернативных) признаков, т.е. при скрещивании брались родительские пары у которых проявлялся либо один, либо другой признак, но не оба вместе. Использовались для опыта генетически чистый материал, т.е. при скрещивании брались организмы которые в ряду поколений не давали расщепления по избранному признаку, т.е. были чистые линии. Проводился тщательный индивидуальный анализ, т.е. прослеживалось наследование признака у каждой особи не только первого поколения, но и наследование признаков в ряде последующих поколений. Использование точного количественного учёта полученного потомства различающихся по контрастным признакам. Это дало возможность установить количественные закономерности в наследовании признаков. Этапы гибридологического метода: 1) Выделение исходных чистых линий. 2) Получение от них гибридов первого поколения. 3) Скрещивание гибридов 1-го поколения между собой и получение гибридов 2-го поколения. 4) Анализ полученных результатов. Успех опытов Менделя объяснялся тем, что изучаемые им признаки контролировались генами расположенными в разных парах хромосом, т.е. контролировались (не сцепленными генами), и Мендель изучал аутосомные хромосомы, т.е. изучал не сцепленные гены.
16. Моногибридное скрещивание и его схема. Изучение наследования признаков в поколении.
С помощью гибридологического метода на основании экспериментального анализа гибридов Мендель установил закономерности наследования признаков в 1865 г. Закономерности: 1) Закон единообразия гибридов 1-го поколения или 1-ый закон Менделя . Он был установлен на основе моногибридного скрещивания «При моногибридном скрещивании все потомки 1-го поколения единообразны и наследуют доминантный признак одного из родителей независимо у какого родителя (отца или матери) был этот признак». 2) Закон расщепления или 2-й закон Менделя : «При скрещивании гибридов 1-го поколения между собой, во 2-ом поколении происходит расщепление в определённом количественном соотношении (Доминантных - 3, рецессивных - 1 признак). Во 2-ом поколении образуется 2-е фенотипические группы расщепления по Ф=3:1 , расщепление по Г=1:2:1 .
17. Строение генов и их влияние на развитие признаков.
Ген Функция гена Ген à Белки à Признаки
Структурные гены В 1961 году оперонная регуляция белка - регулятора . . 2) Оперон содержит функциональные гены : а)Проматор (П) ген б)Ген-Оператор в)Ген-Терминатор (Т)
Ген-Оператор Ген-Регулятор белка-Активатора и белка-Репрессора
18. Основные понятия и термины в современной генетике. Понятие и генотипе и фенотипе. Гомо- и гетерозиготности. Аллельные гены. Доминантность и рецессовность. Генетическая символика.
Доминантность - признак который проявляется в 1-ом поколении и который подавляет развитие другого признака. Рецессивным называется признак не проявляющийся, т.е. подавляемый. Иогансеном был введён в генетику термин ген и он же предложил термин аллельные гены. Аллельные гены - это пара генов, которые располагаются в одних и тех же участках гомологичный хромосом и определяют развитие одного и того же признака. Каждый ген в этой паре называется аллелью . В гаплоидном наборе хромосом имеется только один ген ответственный за развитие данного признака. В соматических клетках содержится 2-а гена. Генотип - совокупность генов, которые определяют развитие наследственных признаков и свойств организма. Фенотип - совокупность признаков и св-в организма, которые сложились в процессе индивидуального развития генотипа. Гомозигота - зигота имеющая одинаковые аллели данного гена, полученные от обоих родителей (оба доминантные или оба рецессивные). Из гомозиготы развивается гомозиготный организм - это организм с одинаковыми аллелями данного гена. Потомство гомозиготной особи однотипна и по фенотипу и по генотипу и не даёт расщепления. Гетерозигота - зигота имеющая 2-е разные аллели данного гена, полученные от обоих родителей (один доминантный, другой рецессивный). Из гетерозиготы развивается гетерозиготный организм - организм имеющий разные аллели одного гена (1-рецессивный, другой доминантный). В потомстве гетерозиготного организма наблюдается расщепление и по генотипу и по фенотипу. Р - родители. ♀ - зеркало Венеры. ♂ - щит и копьё Марса. [АА - гомозигота, аа , Аа - гетерозигота] - аллельные гены. АаВв - дигетерозигота - гетерозигота по 2-ум парам аллелей. Расписать законы Менделя.
19. Генетический код, его основные св-ва.
Генетический кон - это способ записи генетической (наследственной)информации в молекуле ДНК. В молекуле ДНК последовательность нуклеотидов определяет последовательность расположения аминокислот в белковой молекуле. Аминокислоты кодируются 3-мя нуклеотидами , которые кодируют одну аминокислоту. 3-и нуклеотида - это триплет . Триплеты в молекуле РНК называются кодонами , а комплементарные им триплеты молекулы т-РНК - антикоданами . Из 64 триплетов не кодируют аминокислоты три: УАА, УГА, УАГ - триплеты стоп-сигнала . Остальные 61 триплета кодируют аминокислоты. Причём триплет АУГ является стартовым , т.е. с него начинается синтез. Св-ва генетиечского кода : Триплетность. Универсален для всех организмов. Код выражденный. Одна аминокислота может кодироваться несколькими от 1-го до 6-ти кодонами. Только 2-е аминокислоты кодируются одним триплетом: метионин (триплетом АУГ) и триптофан (УГГ). Выраженность кода помогает организму защищаться от вредного воздействия мутации. Неперыкрывающийся, т.е. нуклеотидная послодовательность считывается подряд в одном направлении триплет за триплетом.
20. Современное представление о гене, как единице наследственности.
Серебровский и Дубинин - доказали сложность структуры гена. Ген - участок молекулы ДНК, в котором заложена информация о структуре белка. Ген является единичец наследственной информации. Функция гена : программирование синтеза белков в клетке, но сам ген непосредственно участие в синтезе белка не принимает, т.е. через белки осуществляется контроль за развитие признака. Ген à Белки à Признаки . У низших организмов один ген определяет развитие одного белка, этот белок определяет развитие одного признака. У высших много генов. Один ген определяет развитие многих генов, а эти белки только один признак.
Гены разделяются на 2-е категории : 1)Структурные гены. 2)Функциональные гены. Структурные гены определяют последовательность аминокислот в первичной структуре белка. Функциональные гены контролируют работу структуры генов.
21. Эпистаз. Взаимодействие неаллельных генов. Используя конкретный пример составить схему скрещивания и охарактеризуйте её.
Неаллельными генаминазываются гены локализованные в не гомологичных хромосомах или в разных локусах гомологичных хромосом. При эпистазеген один подавляет действие другого неаллельного гена. Это ген называется: ген-подавитель или ген-супрессор. Расщепление во 2-ом поколении будет в соотношении: 13:3, 12:3:1. По типу эпистаза наследуется: масть у лошадей. Супрессия- это подавление эффекта мутантного гена, восстановление нормы, несмотря на происшедшую до этого мутацию. Рецессивный эпистаз: 9:7, 7/16 комбинаций генотипов особей F2 характеризуются признаком исходных форм скрещивания. У 7 особей из 16 наблюдается рецессивный эпистаз, 3 из них имеют в гомозиготном состоянии ген а и вместе с тем доминантный ген В (ситуация аа>В). Доминантный эпистаз: для него характерен сдвиг числовых показателей расщепления во IIпоколении от классического 9:3:3:1 в сторону 15:1. Так, при скрещивании серых и рыжих лошадей (ССВВ х ссвв) све лошади F1 были серыми, а в F2 из 16 комбинаций 12 оказались серыми, три - вороными и одна - рыжей. Эпистаз наблюдается уже у исходных серых лошадей (С>В) и сохраняется у всех особей F1 (СсВв). Во F2 из 16 комбинаций скрещивания 12 имеют серую окраску, так как у них есть доминантный аллель гена С, 3/16 особей характеризуются вороной мастью - в их генотипе нет доминантного супрессора С. рецессивные гомозиготы ссвв оказываются рыжими, так как повторяют по генотипу исходную форму, взятую для скрещивания. Аналогичная ситуация обнаружена при скрещивании белых и коричневых сибак. Жиыотные Iпоколения были белыми, а во IIпоколении 12 - белые, три - чёрными и одна из 16 комбинаций имела коричневую окраску.
22. Аддитивная полимерия. Её значение в практике животноводства.
При полимерии Признак: А1А1А2А2 а1а1а2а2 . Полимерия 2-х видов: 1)Аддитивная (кумулятивная). 2)Неаддитивная (некумулятивная). При аддитивной полимерии проявление признака зависти от количества доминантных генов в генотипе. Чем больше доминантных генов в генотипе, тем ярче выражение признака. При аддитивной полимерии расщепление во 2-ом поколении будет: 1:4:6:4:1 (Ф=5). При неаддитивной полимерии расщепление не будет зависит от количества доминантных генов. Расщепление во 2-ом поколении будет: 15:1 (Ф=2). Аддитивная полимерия характерна для наследования количественных призанков. Продуктивность животного (удой, масса, настриг шерсти и т.п.), скорость бега и т.д. При аддитивной или дополнительной, полимерии каждый из генов-полигенов вносит значительный вклад в проявление признака.
23. Гаметогенез, сперматогенез и оогенез.
Гаметогенез диффузным Локализованным гонады сперматогенез оогенезом .
В зоне размножения сперматогонии и оогонии содержащие 2 n -хромосом (46 хромосом) сперматоциты и ооциты (содержат 2 n -хромосом ).
В зоне роста сперматоциты 1-го порядка и ооциты 1-го порядка 46 - удвиенных 2 n - хромосом .
В зоне созревания клетки делятся 2-а раза: Мейозом I и II Мейоза I 2-е клетки, сперматоциты 2-го порядка (содержащие n -удвоенных хросомом ) и каждая клетка делится Мейозом II . Образуется 4-е сперматиды (содержащие n -хромосом (23-удвоенных) в сперматозоиды .
В зоне созревания женских половых клеток также 2-а деления Мейоза Iи II. При первом делении образуются 2-е неравные по размерам клетки: одна крупная - это ооцит 2-го порядка (содержит n-удвоенных хромосом), другая маленькая - 1-ое направительное тельце (содержит n-удвоенное). Второе деление Мейзо IIтакже неравное образуется крупная яйцеклетка и 2-ое направительное тельце. 1-ое направительное тельце делится и образуется 2-а вторых редукционных (направленных) телец. Следовательно после оогенеза 3-мелкие направительные и
Оплодотворение
24. Неполное доминирование. Используя пример построить схему скрещивания и охарактеризовать её.
Неполное доминирование проявляется в тех случаях, когда доминантный аллель не полностью подавляет рецессивный аллель и рецессивный аллель частично проявляется. При неполном доминировании в 1-ом поколении проявляют признаки промежуточного между 2-мя родительскими формами. В генетике - это явление известно под названием «промежуточного наследования». При скрещивании гибридов 1-го поколения между собой во 2-ом поколении произойдёт расщепление 1:2:1 (по фенотипу и по генотипу). А - длинная ушная раковина. а - безухие. Р ♀АА(дл) х ♂аа(без). F1 Аа(ср). Р ♀Аа х ♂Аа. F2 АА(дл); Аа(ср); Аа(ср); аа(без). Ф=Г=1:2:1 . Биохимики показали, что в случает промежуточного наследования гомозиготы доминантная и рецессивная производят один и тот же белок. В цитоплазме этот белок суммируется, у гетерозигот (Аа) одинарная доза белка, т.е. гетерозигота производит промежуточную дозу белка и это ведёт к промежуточному выражению признака. (А х А, а х а). Неполное доминирование - это своеобразный эффект взаимодействия аллельных генов, а не простое промежуточное выражение признака. Неполное доминирование может создавать впечатление появления нового признака.
25. Возратное и анализирующее скрещивание, их схемы и значение.
В генетике используется система определённого скрещивания: 1)Возвратным скрещиванием (Беккрос) называется скрещивание гибрида 1-го поколения с одной из родительских форм (сходных по генотипу) гомозиготной по доминантному аллелю. Потомство от этого скрещивания обозначают F в : Р ♀АА х ♂аа. F1 Аа. Р ♀АА х ♂Аа. Возвратное скрещивание используется для преодоления стерильности отделённых гибридов в 1-ом поколении. Все гаметы родительской формы несут доминантную аллель (А), а у гибрида образуются гаметы 2-х видов: (А) и (а), поэтому в результате сочетания этих гамет при оплодотворении в потомстве имеет место расщепление по генотипу 1:1 , а по фенотипу расщепление не происходит. Анализирующее - скрещивание гибрида 1-го поколения с гомозиготой рецессивной: Р ♀АА х ♂аа. F1 Аа. Р ♀Аа х ♂аа. Анализирующее скрещивание используется для определения генотипа гибрида неизвестного происхождения из любого поколения скрещивания. Для анализирующего скрещивания характерно совпадение расщепление по генотипу с расщеплением по фенотипу.
26. Митоз. Его генетическая и биологическая сущность в значении жизни организма.
МИТОЗ (гр. нить) - не прямое деление. Происходит путём сложного деления ядра и цитоплазмы. Митозом делятся соматические клетки, происходит дробление зиготы, образование зародышевых клеток. Митоз составляет часть митотического цикла клетки. Митотический цикл - это существование клетки от одного деления до другого деления. Он состоит из интерфазы и митоза. ДЕЛЕНИЕ ИНТЕРФАЗЫ : G 1 - постмитотический или пресинтетический, S - период синтеза, G 2 - постсинтетический или премитотический. G 1 - период активной жизнедеятельности клетки, клетка очень чувствительна. В этот период входят 5-ть процессов: 1)Накпливается в митохондриях энергия в виде АТФ. 2)Синтез ферментов ДНК полимераза, который принимает участие в репликации молекулы ДНК. 3)В цитоплазме образуются и накаплаваются в ядре 4-е вида нуклеотидов. 4)Синтезируются все виды РНК. 5)Репликация центросом (клеточного центра). S - период. Происходит синтез ДНК, т,е. Удвоение молекулы ДНК. Происходит построение 2-й хроматиды, в которую переходит вновь синтезированная молекула ДНК, образуется двухроматидная хромосома. В этот же пероид синтезируются белки - “гистоны ”. G 2 - период, в котором: 1)Вновь происходит синтез и накопление энергии в виде АТФ. 2)Клетка синтезирует специфические белки, из которых образуются нити веретена деления. МИТОЗ - период деления. Это сложный непрерывный процесс в котором образуются условно 4-и стадии: Профаза, Метафаза, Анафаза, Телофаза. У большинства клеток весь митоз происходит в течении 1-2 часов. ПРОЦЕССЫ В ПРОФАЗЕ : 1)Двухроматидная хромосома спирализуется, она укорачивается и утолщается. Хроматиды связаны вместе при помощи центромер. 2)Хромосомы приближаются к оболочке ядра и ядерная оболочка начинает разрушаться. 3)Ядрышки разрушаются и исчезают. 4)Центриоли расходятся и начинается образование нитей веретена деления. МЕТАФАЗА : 1)Полностью заканчивается формирование веретена деления. Хромосомы при помощи центромер с нитями веретена деления. 2)Двухроматидные хромосомы располагаются на экваторе клетки, т.е. образуют метафазную пластинку. 3)Полностью разрушается ядерная оболочка и кариоплазма и цитоплазма смешиваются. АНАФАЗА : центромеры, которые соединяют 2-е хроматиды делятся и хроматиды движутся к полюсам. ТЕЛОФАЗА : однохроматидные хромосомы деспирализуются, удлиняются и вид тонких нитей. Появляется ядрышко и восстанавливается ядерная оболочка. Генетическая схема митоза : На этом заканчивается кариокинез , т.е. деление ядра и начинается цитокинез,
Каждая хромосома
удваивается и
образуется 2-е пары профаза метафаза анафаза телофаза
хроматид
т.е. деление цитоплазмы. Часть митохондрий, часть комплекса Гольджи, эндоплазм-ой сети и других компонентов цитоплазмы расходятся в дочерние клетки. На экваторе образуется перетяжка и формируются 2-е новые дочерние клетки. МИТОЗ - это наследственно-равное деление. При митозе клетки делятся на 2-е, но не смотря на это число хромосом в дочерних клетках не уменьшается, а остаётся постоянным, причём хромосомы распределяются между дочерними клетками равномерно. БИОЛОГИЧЕСКАЯ СУЩНОСТЬ : равномерное распределение ДНК между дочерними клетками. ГЕНЕТИЧЕСКАЯ СУЩНОСТЬ : сохранение постоянства числа хромосом, в поддержании постоянства кариотипа в поколениях, в образовании генетически-равноценных клеток.
27. Морфология хромосом. Роль центромеры в правильном расхождении хромосом при делении клеток.
Центромера (от лат. centrum, греч. ke`ntrool- средняя точка, центр и греч me`ros- часть, доля), кинетохор, участок хромосомы, контролирующий её движение к разным полюсам клетки во время деления - митоза или мейоза; место прикрепления к хромосоме нитей (микротрубочек) веретена деления. Различают хромосомы с локализованной центромерой (моноцентрические хромосомы) и с диффузной центромерой (голокинетические хромосомы, у которых кинетической функцией обладают, по-видимому, любые участки). Иногда в моноцентрической хромосоме появляется ещё участок с киненетической функцией (неоцентрометрическая активность). Изменение положения центромеры в определённой хромосоме служит критерием выявления хромосомных перестроек. Морфология хромосом изучает многобразие форм хромосом . Ядрышко являются производными хромосом. Хроматин представляет собой плотные структуры в виде гранул, зёрен и образованы молекулами ДНК. Хроматин представляет собой дисперсное состояние хромосом в интерфазе. В процессе деления клеток происходит спирализации молекул ДНК их хроматиновые структуры образуют хромосомы. Термин «хромосома» был предложен немецким учёным Вальдейер в 1888 году . «Chromo» - красный, «Soma» - тело. Хромосомы видны только при делении, хотя они являются постоянными клеточной структуры. Хромосомы - это специализированные клеточные органоиды с помощью которых осуществляется хранение и передача в поколениях генетической информации клетки. Хромосомы лучше всего видны в стадии метафазы митоза. Каждая метафазная хромосома состоит из 2-х продольных копий, которые называются хроматиды . Хроматиды соединяются между собой центромером . Центромера делит хромосому на 2-е части, которые называются плечами хромосом . В зависимости от расположения центромер хромосомы подразделяются: Метацентрические (плечи одинаковые). Субметацентрические (плечи не одинаковые). Акроцентрические (палочковидные). Спутничные (есть перетяжка).
28. Реципрокное скрещивание, их схемы и значение.
Реципрокное (взаимное) - два скрещивания, в которых организмы с доминантными и рецессивными признаками используются и как материнские и как отцовские. 1) Р ♀АА(дом) х ♂аа(рец) - прямое реципрокное скрещивание. 2) Р ♀аа х ♂АА - обратное реципрокное скрещивание. При аутосомном наследовании признаков реципрокное скрещивание результаты будут одинаковыми, т.е. оба родителя в одинаковой степени. Реципрокное скрещивание будет давать разные результаты при наследовании признаков сцепленных с полом. В этом случае наблюдается крест на крест наследование или крисс-кросс. Когда материнский организм передаёт свои признаки сыновьям, а отцовские - дочерям.
Реципрокное скрещивание используется в генетическом анализе для выявления наследственных факторов, локализованных в Х-хромосоме. Кроме того, реципрокное скрещивание позволяют локализовать цитоплазматические наследственные факторы в случае анизогамии, когда в обоих реципрокных скрещиваниях наблюдают передачу потомкам только материнского признака.
29. РНК, виды и роль в наследственности.
В отличии от ДНК РНК состоит из 1-ой цепочки нуклеотидов. В состав нуклеотидов водит 4-е озотистых основания: А, У, Г, Ц. Различают 3-и вида РНК: и-РНК, т-РНК, р-РНК. все виды синтезируются в ядре клетки в присутствии ДНК, затем из ядра они выходят в цитоплазму и там уже функционируют свою определённую функцию. т-РНК находится в цитоплазме и её функция состоит в переносе аминокислот к месту синтеза, т.е. из цитоплазмы к рибосоме. В клетке содержится 20 т-РНК видов. р-РНК содержится в рибосомах и состоит функция в том, что она осуществляет сборку или синтез белка из аминокислот. и-РНК синтезируется на определённом участке молекулы ДНК и является копией этого участка. Транскрипция информации осуществима как с ДНК на РНК, так и обратно. В отличии от транскрипции трансляция представляет собой односторонний переход генетической информации с РНК в структуру полипептидной цепочки белка. обратный процесс не происходит. Некоторые виды (другие) РНК входят в состав белковых нитей веретена деления ядра и т.д. РНК генетическая (наследственная) информация у вирусов, фагов.
30. Нуклеиновые кислоты, ДНК. Строение и роль в наследственности.
В 1869 году шведский учёный Мишер обнаружил в ядре клеток в-ва кислотной природы и их назвал Нуклеиновыми кислотами . Клетки содержат нуклеиновые кислоты 2-х видов: ДНК и РНК. Установлено, что ДНК находятся в хромосомах ядра клетки и её биоллогическая роль состоит в хранении, размножении и передачи наследственной, т.е. генетической информации от родителей к потомкам. ДНК является первичным носителем наследственной информации. Количество ДНК в клетках тела данного типа постоянна . В соматических клетках в 2-а раза больше чем в гаметах. РНК в основном находится в цитоплазме - 90%. Количество в клетке меняется. Нуклеиновые кислоты - это полимеры, мономерами являются нуклеотиды. В состав нуклеотида входит 3-и элемента: 1)Углевод: В РНК - рибоза, в ДНК - дезоксирибоза. 2)Остаток фосфорной кислоты. 3)Озотистые основания : В РНК - А, У, Г, Ц. В ДНК - А, Т, Г, Ц.
Молекула ДНК состоит из 2-х сперальнозакрученных нитей. Эти две нити удерживаются друг с другом с помощью озотистых оснований. А+Г=Т+Ц . Молекула ДНК обладает уникальным свойством способностью к идентичному удвоению, т.е. репликации или самоудвоению . Перед самоудвоением благодаря ферменту происходит раскручивание 2-х спиралей. Происходит разделение её цепей между озотистыми основаниями. Построение 2-й цепочки молекулы ДНК происходит по типу матричного процесса, т.е. новые молекулы синтезируются в точном соответствии с химической структуры уже существующих молекул. вО время репликации ДНК каждая из 2-х её цепей служит матрицей для образования новой цепи. Репликация ДНК протикает быстро. Напритмер 900 нуклеотидов в минуту.
31. Балансовая теория определения пола. Паталогия по половым признакам.
В 1922 году американским генетиком Бриджисом была разработана «Балансовая теория определения особей ». Пол особей определяется соотношением числа половых хромосом и аутосом в зиготе (их балансом). Он проводил опыты над Дрозофилами. В опытах на мухах дрозофилах были получены потомки имеющие признаки того и жругого пола. И были получены интерсексы, т.е. особи с промежуточным развитием пола. Среди мух дрозофил были особи с сильновыраженными половыми железами: сверхсамки и сверхсамцы. Цитологический анализ таких необычных особей у дрозофилы показал, что для их кариотипа характерна и необычное соотношение половых Х-хромосом и аутосом. Оказалось повышенная доля аутосом по отношению к Х-хромосомам приводит к усилению мужских признаков, а при повышении доли Х-Хромосом усиливается появление женских признаков. Определение пола особей зависит от полового индекса: Х:А .
Если половой индекс = 1, то рождаются нормальные самки: 2Х:2А=1♀ нормальные . Если половой индекс = 0,5, то рождаются нормальные самцы: Х:2А=0,5♂нормальные . Если половой индекс больше 1, то рождаются сверхсамки: 3Х:2А=1,5 сверх♀ . Если половой индекс меньше 0,5, то рождаются сверхсамцы: Х:3А=0,33 сверх♂ . От 1 до 0,5 - интерсексы - имеющие (особи) промежуточные признаки ♀ и ♂: 2Х:3А=0,66 (интерсексы) .
У ряда животных (различных видов), а также и у человека обнаружена патология по числу половых хромосом . Основной причиной таких ненормалей является нерасхождение половых хромосом при делении, при этом число аутосом ненарушается (в норме). У самок млекопитающих в результате нарушения в расхождении 2-х Х-хромосом образуются не нормальные гаметы. Не нормальные гаметы выражается в числе половых хромосом, т.е. одна гамета содержит 2Х хромосомы, а другая - неодной Х-хромосомы. А в результате оплодотворения ненормальных гамет с нормальными гаметами образуется 4-е типа зигот: ХХХ; ХО. ХО - синдром Тернера - ♀ . ХХ Y - синдром Клайнфельтера - ♂ .
XXY. Генетическими наследованиями в 1949 году и исследованиями в дальнейшем установлено, что нормальные соматические клетки мужских и женских особей характеризуется наличием или отсутсвием в них небольшого хроматинового тельца, который обнаружен при окраски. Это хроматиновое тельце называется тельце Барра или половой хроматин . Число телец Барра всегда на единицу меньше числа Х-хромосом.
32. Полимерия не аддитивная, её значение в практике животноводства.
При полимерии признак находится под контролем нескольких пар неаллельных генов, которые однозначно влияют на этот признак. Признак: А1А1А2А2 - 2-е пары неаллельных генов (А1а1а2а2; А1А1А2а2). а1а1а2а2 . Полимерия 2-х видов: 1)Аддитивная (кумулятивная). 2)Неаддитивная (некумулятивная) . При аддитивной полимерии проявление признака зависти от количества доминантных генов в генотипе. Чем больше доминантных генов в генотипе, тем ярче выражение признака. При аддитивной полимерии расщепление во 2-ом поколении будет: 1:4:6:4:1 (Ф=5). При неаддитивной полимерии расщепление не будет зависит от количества доминантных генов. Расщепление во 2-ом поколении будет: 15:1 (Ф=2). Некумулятивная полимерия характеризуется тем, что для полной выраженности признака достаточно доминантного аллеля одного из полимерных генов (олигогена). При неаддитивной полимерии особи сходны кроме рецессивной аавв . Неаддитивная полимерия была обнаружена при скрещивании растений пастушьей сумки с треугольными и овальными струкчами. У потомства Iпоколения все стручки треугольные, а во IIпоколении 1/16 растений имеет овальные стручки, у остальных же, как и в Iпоколении, они треугольные.
33. Типы хромосомного определения пола у животных: сингамное, эпигамное, прогамное.
1)Сингамный тип - происходит в момент оплодотворения. Такой тип характерен для млекопитающих, человека, мухи дрозофилы, моллюсков и иглокожих, для большинства амфибий и рептилий. У этих животных самцы являются носителями неодинаковых половых хромосом и при гаметтогенезе образуют сперматозоиды 2-х типов (гетерогаметны-самцы). У самок обе половые хромосомы одинаковы и при гаметогенезе образуеюся гаметы одного типа (самки -
Определяется в ходе гаметогенеза у родительской формы. Птицы, рыбы, амфибии и рептилии, насекомые- чашуекрылых. У эти животных женский пол гетерогаметен (ХY), а мужской гомогаметен (ХХ), т.е. ♂ - одного вида гамет, а у ♀- двух видов гамет образуется. 3)Эпигамный тип - пол определяется в процессе онтогенеза. Характерен для пчёл, муравьёв, наездников. У них нет половых хромосом. Только одни аутосомы. Пол особи определяется числом, т.е. кратностью числа аутосом. При диплоидном наборе хромосом 32 пчёл из оплодотворённой яйцеклетки при гаплоидном наборе хромосом развиваются трутни. У них нет отца. 4)ХХ - ХО тип - харектерен для клопов, кузнечиков. У них гетерогаметны - мужской пол, но из половых хромосом но содержит только одну Х-хромосому (ХО), а самки - гомогаметны (ХХ).
34. Комплементарное взаимодействие неаллельных генов. Используя конкретный пример, составить схему скрещивания и охарактеризуйте её.
Неаллельными генами называются гены локализованные в не гомологичных хромосомах или в разных локусах гомологичных хромосом. При комплементарности 2-а неаллельных гена находясь вместе в одном генотипе определяют развитие нового признака. При комплементарном взаимодействии неаллельных генов во 2-ом поколении при скрещивании гибридов 1-го поколения между собой возможны следующие расщепления: 9:7, 9:6:1, 9:4:3, 9:3:3:1, как при дигибридном скрещивании, но в отличие от дигибридного скрещивания по 2-ум признакам при комплементарном скрещивании идёт только по одному признаку. Пример форма гребня у кур : дом.ген R. - розовидная форма.
35. Типы действия генов: плейотропия и модифицирующее действие.
Плейотропия - множественное действие генов, при котором один и тот же ген определяет развитие нескольких признаков. Примеров много: у человека один и тот же ген определяет цвет волос (рыжий), веснушки. Плейотропия связана с тем, что такие гены контролируют синтез белков-ферментов, которые принимают участие в нескольких биохимических обменных реакций в клетке и организме и тем самым оказывают влияние на развитие нескольких признаков, причём несвязанных между собой. Практически каждый ген контролирует определённый этап метаболизма. Многоэталонность метаболизма приводит к тому, что нарушение метаболизм на одном этапе неизбежно сказывается на признаках. Часто плейотропные гены вызывают видимые внешние морфологические изменения, но в внутренние строение внутренних органов; и в итоге сказываются на жизнеспособность или привести организм к гибели. Гены приводящие организм к гибели на определённой стадии онтогенеза смерти называют летальными генами. Летальные гены - обычные гены, от других они отличаются тем, что своё воздействие через ферменты оказывают на развитие и функционирование жизненно важных органов изменяя их так к понижению жизнеспособности этих органов. Часто летальные гены обладают двойным действием: с одной стороны вызывают развитие полезного для человека хозяйственно-полезного признака (окраска животных). С другой стороны со стороны плейотропии снижает жизнеспособность и даже приводит к гибели. Среди овец каракульской породы встречаются животные с красивой серой серебристой окраской (Аа ûАа). F 1 = (АА; Аа; Аа;)сер. мех. Аа чёрный мех. АА (25%) - погибают при переходе на питание сухим кормом. Ген - доминантный вызывает серую окраску у овец и этот же ген оказывает влияние на развитие желудочно-кишечного тракта. У гомозиготных больных овец был очень узкий сфинктер, поэтому ягнята - молоком живые были, а травой - погибали. Было запрещено скрещивать серых овец с серыми.
Примером плейотропного действия гена у человека служит рецессивная мутация в гене, которая контролирует синтез в полипептидной цепи в молекуле гемоглобина. Мутация вызывает замену одной аминокислоты и это влечёт в изменении морфологии эритроцитов (серповидной формы), к нарушению сердечно-сосудистой, нервной систем и т.д. Гомозиготность приводит по этой мутации к гибели. Часто летальные гены являются рецессивными, поэтому гетерозиготы нормальны по фенотипу, а у рецессивных гомозигот они проявляются. Летальные гены встречаются у животных всех видов в аутосомах и половых хромосомах. К настоящему времени выявлено у КРС 46 летальных генов; кур - 45; Лошадь - 10; Свинья - 18; Овца - 15.
Модифицирующее действие генов: признак под контролем ßгена модификатора (ген не имеет фенотипического выражения); основного. Пример: Модифицирующего действия белая пятнистость шерсти у КРС. Ген основного действия вызывает образование белых пятен на голове, туловище и т.д., а один ген модификатор влияет на размер белых пятен, а другой ген модификатор оказывает на распространение этих пятен. Гены модификаторы обнаружены у пушных зверей. У кроликов усиливал голубую масть у Венской голубой породы. У каракульских овец: модификационный ген доминантный - ослабитель основного окраса (черного или коричневого). Установлено три пары генов модификационной влияющие на количество красного пигмента в волосе КРС сементальской породы. У гомозиготных по рецессивному гену красной масти животных интенсивности окраски колеблется от вишнёвого до палевого.
36. Задачи современной генетики. Значение генетики для теории и практики животноводства и растений.
Биосинтез белка является прямой функцией генов. Без синтеза же белков невозможны жизненные процессы, поэтому решающее значение генетики среди других биологических наук очевидно. Гены определяют не только особенности признаков, но и возможность их развития, тем самым возможность развития и существования клетки, организма. В то же время современная генетика обязана своими достяжениями развитию многих наук: физики, химии, кибернетики, математики, цитологии, эмбриологии, молекулярной биологии и др. достяжения генетики вносят вклад в общую теорию развития, обогащают конкреьным содержанием диалектико-материалистическое мировоззрение. Однако развитие промышленности, использование двигателей внутреннего сгорания, химических веществ для борьбы с сорняками и вредителями в сельском хозяйстве порою загрязняют окружающую среду вредными соединениями, нередко и такими, которые могут вызывать нежелатеные мутации. Изменение среды обитания сопровождается изменениями в ней характера биологических связей, нарушением баланса этих связей, что в итоге может привести к гибели системы биологических связей. Использование теоретических и практических основ генетики позволяет выводить не только сорта растений, он и породы животных, устойчивых к возбудителям болезней и неблагоприятным факторам внешней среды, породы, отличающиеся более высокой продуктивностью. Задача (в ветеринарии) при решении создании эффективных противомикробных и противовирусных вакцин, а также продуцентов противовирусного препарата - интерферона.Составление генетических карт являются важной задачей современной генетики. Карта хромосом даёт представление о том какие гены наследуются независимо друг от друга, а какие сцеплено. Как часто между ними происходит кроссинговер. Создание генетических карт позволяет планировать работу по получению организмов с определёнными сочетаниями признаков.
37. Сущность закона гомологичных рядов наследственной изменчивости Вавилова.
Ещё в 1920 году Вавилов сфомулировал закон гомологических рядов в наследственной изменчивости у близких видов и родов, который биологи сравнивают с периодической системой Д.И. Менделева. Вавилов обнаружил, что «1.Виды и роды генетически близкие между собой, характеризуются тождественными рядами наследственной изменчивости с такой правильностью, что зная ряд форм для одного вида, можно предвидеть нахождение тождественных форм у других видов и родов. Чем ближе генетически расположены в общей сисмете роды и линнеоны, тем полнее тождество в рядах их изменчивости. 2.Целые семейства растений в общем характеризуются определённым циклом изменчивости, проходящей через все роды, составляющие семейство». Так, у мягкой пшеницы есть растения с остистыми, безоститыми, полуостистыми колосьями; белоколосые, красноколосые, черноколосые, сероколосые формы и т.д. Родственные мягкой пшенице виды имеют те же формы. Вавилов объяснил это тем, что у родственных организмов изменчивость признаков идёт в одном направлении - параллельно. Такую параллельную изменчивость имеют виды не только одного рода, но и близких по своему происхождению рода, но и близких по своему происхождению родов, например, пшеницы, ячменя, ржи и других злаков. Этот закон помогает учёным ориентироваться среди огромного разнообразия живых существ. Он облегчает поиск нужных для селекции хозяйственных признаков растений и животных.
Руководствуясь законом гомологических рядов, учёные нашли, например, негорькие, съедобные для пастбищных животных формы люпина, одновременно обогощающие почву азотом, которые стали родоначальником новой, кормовой формы люпина.
38. Основные положения хромосомной теории наследственности Томаса Моргана.
Положения : Гены находятся в хромосомах. Каждый ген в хромосоме занимает определённое место (локус ). Гены в хромосомах расположены в определённой линейной последовательности. Каждая хромосома представляет собой группу сцепления генов. Число групп сцепления у каждого вида равно наследуются сцепленно. Признаки, гены которых в одной хромосоме наследуются сцепленно. Новые сочетания генов находящихся в одной хромосоме возникают в результате кроссинговера в процессе мейоза. Расстояние между генами в хромосоме пропорциональна процессу кроссинговера между ними.
39. Сцепленное наследование. Используя конкретный пример, составить схему скрещивания и охарактеризуйте его.
Например
40. Понятие оь изменчивости. Виды изменчивости: модификационная изменчивость.
Изменчивость существует 2-х видов: 1)Ненаследственная изменчивость или фенотипическая: модификационная изменчивость. 2)Наследственная или генотипическая изменчивость: а)Коррелятивная. б)Комбинативная. в)Мутационная.
Модификационной изменчивостью называют разнообразие фенотипов вызванная факторами снешней среды. Особенности модификационной изменчивости: Она не связана с изменениями гена, генотипа и поэтому по наследству не передаётся. Она носит групповой характер, т.е. наблюдаются сходные изменения признака у всех особей данного вида. Она имеет приспособительное значение и можно её предсказать. Например животных жившых на равнинах переселить в горы, то у них у всех повысится поддержание гемоглобина в крови. Она имеет приделы. Этот предел называется «нормой реакций ». Норму реакции обуславливают гены организма, а от среды заисит какой вариант в пределах этой нормы реакции реализуется в данном случае и модификационная изменчивость изображается графически. Вариационная кривая модификации отражает частоту встречаемости различных форм. Чаще всего встречаются средние показатели вариационного ряда, а особи с минимальным или максимальным развития признака бывают немного. Модификационная изменчивость имеет практическое значение. Создание оптимальных условий приводит к полной реализации данного генотипа. Изменяя условия среды можно раскрыть границы нормы реакции её генов и это даст возможность проводить селекцию хозяйственнополезных признаков при определённых условиях среды.
Генотипическая изменчивость - она связана с изменением генотипа. а)Коррелятивная (соотносительная) изменчивость . При этой изменчивости изменение одних признаков приводит к изменению других. Например : с увеличением живой массы коров первотёлок, возрастает удой - это положительная корреляция, а с увеличением продуктивности уменьшается содержание жира в молоке - отрицательная корреляция. Факторы, которые определяют коррелятивную изменчивость - это плейотропное дейтсвие генов и взаимосвязь между признаками. б)Комбинативная связана с получением новых сочетаний (комбинаций генов в генотипе). Сами гены при этом не изменяются, но новые сочетания их между собой приводят к появлению организмов с новыми фенотипами. Комбинативная изменчивость наблюдается в результате комплементарного взаимодействия неаллельных генов (форма гребня). Комбинативная изменчивость лежит в основе создания новых форм при селекции с/х животных. Например : Орловская рысистая порода Лошадей получена комбинацией в скрещивании жеребца Арабской верховой породы с кобылами. Голландских и Дадских пород. в)Мутационная изменчивость Мутация Мутации мутационной теории были изложены Де-Фризом.
мутагенезом мутантом спантанными индуцированные мутации .
41. Мутационная изменчивость. Основные положения мутационной изменчивости.
Мутационная изменчивость - это изменчивость, которая связана с изменением наследственного материала. Термин «Мутация » впервые был предложен голландским ботаником Де-Фризом в 1901 году. Мутации - это внезапно-возникающие генетические изменения св-в и признаков. Основы мутационной теории были изложены Де-Фризом. Основные положения мутационной теории : Мутации возникают внезапно и возникают у единичных особей. Мутации носят случайный ненаправленный характер. Мутации являются изменением наследственных (генетических) структур в соматических и половых клетках. Мутации передаются из поколения в поколение. Мутации происходят в разных направлениях. Они могут быть вредными, нейтральными и полезными для организма.
Более 90% мутаций оказывают влияние на жизнедеятельность организма. Процесс возникновения мутаций называется мутагенезом . Организм у которых произошла мутация называют мутантом . Мутации могут возникать в природе случайно без вмешательства человека и их называют спантанными . Мутации могут быть вызваны искусственно путём воздействия различных мутагенов - индуцированные мутации .
42. Кроссинговер, как причина неполного сцепления генов.
Сцепление может быть полным и неполным .
Гибриды которые образуются в результате слияния кроссоверных гамет называются кроссоверными особями. И кроссоверных особей в потомстве около 17% . Результаты многочисленных опытов указывают, что гены расположенные в одной хромосоме действительно сцеплены и лишь с определённой частотой разъединяются в следствии кроссинговера. Частота кроссинговера марганиды или санти-морган Генетической картой хромосом Морганиды
43. Сцепление генов в хромосоме. Группы сцепления.
Для каждого вида характерна определённое и постоянное число хромосом. Количество признаков и генов которые их контролируют значительно больше, чем число хромосом. Например : у дрозофилы известно около 1000 генов, а всего хромосом 4-е пары. У человека несколько 1000 генов и всего 23 пары хромосом. Гены находящиеся в одной хромосоме называются «сцепленными» и образуют «группу сцепления», т.е. они (гены) передаются вместе (сцепленно) с начало в гамету, а затем следующему поколению. Число групп сцепления равна гаплоидному числу хромосом . При изучении наследования сцепленных генов запись генотипов видоизменяется. При независимом наследовании признаков АаВв, при зависимом сцепленном наследовании: АВ//ав. АаВв А | | а В | | в АВ//ав А | | а
Сцепление может быть полным и неполным .
Гибриды которые образуются в результате слияния кроссоверных гамет называются кроссоверными особями. И кроссоверных особей в потомстве около 17%.
44. Спантанные и индуцированные мутации. Мутагенные факторы. Частота и причины мутаций.
Индуцированные мутации возникают в результате вызванного действием мутагенов нарушения нормальных процессов репликации, рекомбинации, репарации, формации. Спонтанные мутации возникают как ошибки при воспроизведении генетического материала, поскольку редупликация не происходит с абсолютной точностью, а процессы репарации не обладают абсолютной эффективностью частота спонтанного мутирования у каждого вида генетически обусловлена и поддерживается на оптимальном уровне.
: 2)Хромосомные мутации 3)Геномные мутации полиплоидия Гетероплоидия Гетероплоидия
45. Генетические карты хромосом. Линейное расположение генов в хромосоме.
Результаты многочисленных опытов указывают, что гены расположенные в одной хромосоме действительно сцеплены и лишь с определённой частотой разъединяются в следствии кроссинговера. Частота кроссинговера зависит от расстояния между генами. Чем ближе расположены гены в хромосоме, тем реже они разъдиняются при перекрёсте и наоборот. Частотта кроссинговера свидетельствует о линейном расположении генов вдоль хромосомы, т.е. гены располагаются в один ряд, в определённых местах и на определённых расстояниях друг от друга. О расстоянии между генами судят по частоте кроссинговера. За единицу измерения генетического расстояния между генами в хромосоме принят 1% перекрёст, который получил название: марганиды или санти-морган равный проценту кроссинговера между ними. Зная расстояние между генами можно построить карты хромосом. Генетической картой хромосом называют схему взаимного расположения генов находящихся в одной группе сцепления. Морганиды - это генетическое расстояние между генами. Расстояние между генами соответствует частоте кроссинговера. Частота кросиинговера выражается в % и вычисляется по формуле: %=(число кроссоверов/общее число особей)*100% . Составление генетических карт являются важной задачей современной генетики. Карта хромосом даёт представление о том какие гены наследуются независимо друг от друга, а какие сцеплено. Как часто между ними происходит кроссинговер. Создание генетических карт позволяет планировать работу по получению организмов с определёнными сочетаниями признаков.
46. Понятие о популяциях и чистых линиях. Опыты Иогансена. Генетика популяций, формула и закон Харди-Вайнберга.
Вид состоит из популяций. Популяция - это совокупность особей одного вида, населяющих определённую территорию и обменивающихся генетическим материалом. Существуют природные и искуственные популяции. Породы житвотных являются популяциями созданные человеком (созданные искусственным путём). Стадо какой-то породы является популяцией. Популяцию составляют генетически не одинаковые особи. Совокупность генов свойственная всей популяции называется генофондом данной популяции. «Чистая линия » используется для обозначения группы организмов однородных по генотипу. Датский учёный Иогансен проводил исследования по выявлению эффективности отборов в популяций и чистых линиях. Он проводил опыты на самоопыляющемся растении фасоли. В течении 6-ти лет от проводил отбор фасоли по массе семян. В первом опыте Иогансен собирал семена со многих разных растений. Они оказались разными по массе. Биометрическая обработка показателя массы дала варицаонную кривую. Семена с максимальным и
в среднем мелкие по 0,3-0,4 гр., но крупные родительских. С растений выращенных из средних по массе семян собрали средние же. и результаты в течении 6-ти лет были одинаковы.
Во втором опыте Иогансен собирал семена отдельно с каждого растения фасоли и они также оказались разными по массе. Далее крупные 0,7 семена, отобрал и мелкие 0,2 и посеил их отельно и со всех вырасщеных из них растений получил в среднем одинаковые средние фасолины массой 0,5гр. В течении 6-ти лет - вывод тот же самое, что в первом опыте . Во втором случае, когда он брал семена с одного растения всё потомство было одинаковым по генотипу, потомство одного растения Иогансен назвал «чистой линией ». В генетическом отношении они были однородны., хотя были несходны по генотипу. Фенотипические различия между такими особыми, т.е. измечивость в пределах чистой линии связано не с их различием по генотипу, а с воздействиями среды. Эти различия являются модификационными и потомству не передаётся.
На основе результатов 2-го опыта Иогансен установил, что отбор модификаций в пределах чистой линии, где особи генетически однородны недаёт положительного результата. В 1-ом случае когда семена с разных растений отбор был эффективен, т.к. они были наследственно неоднородными, т.е. различались по генотипу. Группу наследственно-разнородных особей одного вида Иогансен назвал популяцию . Изменчивость в пределах популяции обусловлена генотипическими различиями. В популяции благодаря разной наследственности отбор эффективен.
Иогансен изучая изменчивость количесвтенных признаков впервые показал наличие 2-х типов изменчивости: генетическую и не наследственную. Было показано, что у растений из одного исходного организма отобранного из популяции можно создать чистую линию, а из неё новый сорт. Изменчивость в пределах которого будет модификационным (ненаследственной), а исходная наследственность будет сохраняться в нескольких поколениях. Таким образом будет сохраняться в нескольких поколениях. Таким образом установление эффективности влияния отбора в популяции и отсутствие эффекта отбора в чистых линиях имеют большое значение для селекционной практики.
Популяция характеризуется определённой генетической структурой, т.е. определённым соотношением генотипов. Если в популяции происходят свободные скрещивания ♀ и ♂ любого генотипа, то такая свободноскрещивающаяся популяция находится в рвновесии, т.е. в ней сохраняется определённое соотношение генотипа. В 1908 году английский математик Харди и немецкий врач Вайнберг определили структуру популяции по генотипам. Предложили формулу для определения этой структуры. И сформулировали закон о структуре свободноразмножающихся особей. Аа х Аа. р - это частота встречаемости доминантного гена А. q - это частота встречаемости рецессивного гена а . При скрещивании
характризуется 2-мя параметрами : 1)Частота встречаемости генов. 2)Частота встречаемости генотипов. Под генетической структурой популяции понимается частоты встречаемости генов и частоты встречаемости генотипов, которые выражаются в % или долях единицы. Пример : В стаде КРС 84% черной масти животных - р 2: 2рq. 16% красной масти животных (q 2) рецессивный признак. q 2 aa=16%. qaa=4=0,4. рA+qa=1. рA=1-0,4=0,6. р 2 AA=36%:48%(Aa):16%(aa). Генетическая структура популяции не изменяется от поколения к поколению, т.е. находится в состоянии генетического равновесия при определённых условиях : Если размеры популяции велики. Если спаривание происходит случайным образом. все генотипы одинаково плодовиты. Новых мутаций не возникает. Отсутсвует обмен генами с другими популяциями.
47. Факторы влияющие на изменение генетической структуры популяции.
Генетической динамикой популяции называется изменение соотношения частот генов и частот генотипов в поколении. Популяция обладает и генетической пластичностью (изменяется генетическая структура) и обладает генетическим гомеостазом (постоянством). Эта 2-а св-ва популяции обеспечивают её генетическую динамику. Причинами изменения генетической структуры популяции могут быть ряд факторов : Мутационный процесс. Отбор естественный и искусственный. Миграция особей из или в популяцию. малочисленность особей в популяции вызывая дрейф генов.
Мутационный процесс : мутации - это внезапные изменения наследственного материала. Впервые факт насыщения популяции мутациями установил советский учёный Четвериков С.С. в 1926 году. Путём мутации гены изменяются. Каждый отдельный ген мутирует очень редко, но в популяции огромное число генов, поэтому и мутирует большое число генов. Появление нового мутантного гена вызвает изменения чистаты исходного гена. Мутантные гены могут оказывать разное влияние на популяцию, это зависит от: Характера изменений, которые вызываются мутантными аллелями. Характера их действия. Мутации в популяции часто возникают в рецессивной форме и проявляются в гомозиготном состоянии, а в гетерозиготном состоянии - это скрытые генетические изменения. Наличие и распространение в популяции скрытых рецессивов называется «Генетическим грузом ». При разведении с/х животных генетический груз создаёт условия для породообразования. Например : а Англии выведено на базе мутантных котортконогих овец Анконская порода. В пушном звероводстве и кролиководстве распространяются и поддерживаются селекцией альбиносные животные.
Отбор : отбор не создаёт новых генов, но через устранения от размножения особей меняет частоты соотношения генов и частота встречаемости генотипов. Естественный отбор действуя в природной популяции способствует повышению приспособленности организмов к различным условиям среды. Естественный отбор сохраняя одни генотипы и устраняя другие приводит к разнообразию организмов в популяции. Чем выше приспособленность у особей данного генотипа, тем выше частота распространения генотипа в популяции. Чем лучше приспособлен организм, тем больше выживаемость, большая плодовитость и скорость развития. Естественный отбор затрачивает жизненноважные признаки особей в природных популяциях. Искусственным отбором называется любая человеческая деятельность в результате которой одни организмы получают преимущество над другими. Искусственный отбор может способствовать сохранению в популяции особей с определёнными аллелями и генотипами, т.е. отбор может идти в пользу аллелей и наоборот отбор может идти на устранение данного аллеля. Отборы в пользу: доминантного аллеля, рецессивного аллеля, гетерозиготы, против гетерозиготы. Стабилизирующий отбор направлен на поддержание в популяции среднего значения признака и устраняет особей которые отклоняются от среднего значения признака. Учение о стабилизирующем отборе разработал советский учёный Шмальгаузен И. Стабилизирующий отбор уничтожает крайние отклонения стабилизирует среднюю норму выраженности признака, при этом наступает стадилизация генетической изменчивости и частоты генов преобретают равновесное состояние. Стабилизирующий отбор характеризуется тем, что на протяжении ряда поколений формируются наиболее приспособленные фенотипы и популяция достигает высокого уровня приспособленности. Пример : наблюдение сильных ветров, уроганов (погибли воробьи). Движущий (направленный) отбор (Дарвин). При медленных и неуклонно изменяющихся условиях внешней среды происходит сдвиг средней нормы в ту или иную сторону. Отбор в таких случаях благоприятствует тем особям, у которых появляются приспособления к новым суловиям окружающей среды. При искусственном направлении отбора происходит постепенный сдвиг среднего значения признака в сторону повышения или понижения в зависимости от целей (условий) селекций.
Миграция : у поступление в популяции новых её членов из другой популяции или наоброт (продали). Миграционные процессы наблюдаются при использовании искусственного осеменения. Генетическая структура популяции может изменяться даже если не будет влиять отбор миграции и мутации. На генетическую структуру популяции оказывает влияние её численность. В изолированных популяциях, при малой численности (менее 500) особей наблюдается явление известное под названием «генетика автоматический процесс » или «дрейф генов » - это изменение частот генов и генотипов под влиянием случайных причин. В небольших популяциях случайные события (преждевременная гибель особи), которая была единственным обладателем какого-то аллеля приведут к исчезновению этого аллеля в популяции. Например : если данный аллель встречается в популяции из 1 млн. особей, с частотой 1%, то им будут обладать 10 Тыс. особей, а если в популяции из 100 особей - этот аллель будет иметься только у одной особи. Поэтому случайная утрата этой особи влечёт за исчезновением единственного аллеля. В популяциях ограниченного размера нельзя избежать близкородственных скрещиваний - явление «Инбридинга» - это состояние, которое приводит к состоянию увеливению гомозиготности, к снижению воспроизводительных функций особей; снижение жизнеспособности и к выражденею и к гибели.
В популяции фенотипическое разнообразие формируется не одинаково при наследовании качественных и количесвтенных признаков. Это зависит от : Количества генов, которые принимают участие в формировании качественных признаков и количественных (число). При формировании качественных признаков (масть) участвует мало генов (либо 1-а, либо 2-е пары), а при количественных признаков - множество генов (почти весь генотип). Эффекта отдельного гена. Взаимодействия генов. Формирование качественных признаков (участвует 1 или 2-а вида взаимодействия генов). Количественных признаков (участвует все видыв сумме взаимодействия генов). Разнообразия среды на формирование признаков (фенотипов). Качественные признаки (среда не влияет). Количественные признаки (среда оказывает влияние на фенотипов как прямое так и косвенное).
48. Наследование признаков сцепленных с полом.
При аутосомном наследовании оба родителя в одинаковой степени передают свои признаки в одинаковом количестве потомству. Наследование признаков, гены которых находятся в Х-хромосоме называют «наследование сцепленное с полом ». Генетическими исследованиями установлено, что Y-хромосома, она генетически энертна (она наследственно пуста). При наследовании признаков сцепленных с полом оба родителя не в одинаковой степени способны передавать свои признаки потомству. Это явление было открыто Г. Мограном и его сотрудниками в исследованиях на дрозофиле. Они обнаружили, что у дрозофилы окраска глаз является признаком сцепленным с полом. А - красные глаза (доминантность). а - белые глаза (рецессивность). При полном скрещивании : Р ♀Х А Х А х ♂Х а Y. F1 Х А Х а (кр); Х А Y(кр). Р ♀Х А Х а х ♂Х А Y. F2 Х А Х А; Х А Y; Х А Х а; Х а Y. При обратном скрещивании : Р ♀Х а Х а (бел) х ♂Х А Y(кр). F1 Х А Х а; Х а Y. F2 Х А Х а; Х а Х а; Х А Y; Х а Y.
При анализе наследования признаков сцепленных с полом было установлено, что половые хромосомы гомогаметного материнского организма передают признаки сыновьям, а единственная Х-хромосомя гетерогаметная мужского пола передаётся дочерям, т.е. признаки наследуются крест-накрест . У человека также известен ряд признаков (60 генов в Х-хромосоме): дальтонизм, гемофилия, полидоктения (6 пальцев). Эти признаки - рецессивных генов. У кур, рыб гетерогаметным полом является самки, а гомогаментным самцы. У кур наследуется сцепленно с полом по типу крест-накрест (попересно-полосатая окраска).
49. Признаки ограниченные полом и их наследования.
Ограниченным полом называются те признаки, которые свойственны только одному какому-то полу, несмотря на то, что гены обуславливающие данный признак присутствуют у обоих родителей. Такие гены обычно локализованы в любой паре аутосом (они не сцеплены с полом), поэтому и ♀ и ♂ передают их потомству в равной степени.
У человека облвсение со лба типично для мужчин (признак ограниченный полом). Ген определяющий облысение локализован в аутосоме и у мужчин этот ген ведёт себя как доминантный из-за присутствия мужских половых гормонов, у женских он ведёт как рецессивный ген. Способность коров давать молоко. Были одинаково с коровами влеяют на продуктивные качества потомства, т.е. на молочную продуктивность, т.е. эти признаки передаются как через мать, так и через отца. Бык несёт гены определяющие высокую молочность. Петухи одинаково с курами вляют на яйценоскость кур.
50. Классификация мутаций по генотипу. Генный, хромосомный и геномный и их характеристика.
По характеру изменения генотипа выделяют типы мутаций : 1)Генные или точковые мутации - это наиболее часто встречающиеся. Они связаны с изменением последовательности нуклеотидов в молекуле ДНК. Они приводят к тому, что мутантный ген перестаёт работать и тогда или не сентезируется информационная РНК и белок или синтезируется белок с изменённым составом, а следовательно и с свойтвом.они приводят к разнообразным морфологическим, биологическим, физиологическим изменениям. Мутировать могут все гены, которые контролируют развитие любого признака. Большинтсво генных мутаций являются вредными для организма. В основе многих наследственных заболеваний лежат генные мутации такие как гемофилия, альбинизм, дальтонизм. 2)Хромосомные мутации - это перестройки хромосом (хромосомные аберрации). Различают несколько типов хромосомных аберраций: Дупликация - это удвоение участка. Делеция - это нехватка участка (АВСДàАСД). Инверция - это поворот участка (АВСàАСВ). Транслокация - это обмен участками между не гомологичеными хромосомами (АВСàАКМ). Хромосомные мутации возникают в результате отклонений в нормальном процессе клеточного деления. Когда происходит развитие хромосом, хроматид и воссоединение в новых сочетаниях. 3)Геномные мутации , которые приводят к изменению числа хромосом. Наиболее распространённым является полиплоидия - это кратное изменение числа хромосом (3n- триплоид; 4n- тетраплоид и т.д.). Полиплоидия широко распространена среди растений. Возникновение полиплоидии связано с нарушением мейоза или митоза. У животных полиплоидия редко, связана с тем, что хромосомный баланс определения пола характерен для животных препятствуют для полиплоидии. Гетероплоидия - это геномная мутация, которая заключается в изменении числа хромосом не кратная гаплоидному (2n±1). Гетероплоидия возникает при нарушении в расхождении хромосом в результате деления клеток. Люди имеющие трисомию (47 хромосом) имеют признаки болезни дайна (маленький: нос, голова, слабое развитие). Даун чаще всего у детей рождённые не молодыми родителями. У КРС трисомия тоже встречается и наблюдаются морфологические изменения (укорочение нижней челюсти; карликовость; водянка).
51. Теория Жакоба и Моно и регуляции синтеза и-РНК и белков.
Гены разделяются на 2-е категории : 1)Структурные гены. 2)Функциональные гены. Структурные гены определяют последовательность аминокислот в первичной структуре белка. Функциональные гены контролируют работу структуры генов. В 1961 году французские учёные Жакобо и Вано разработали теорию регуляции активности генов (это включения или выключения тех или иных генов в процессе синтеза белков на уровне транскрипции) - оперонная регуляция . Оперон - участок молекулы ДНК, на котором осуществляется трунскрипция молекулы и-РНК под контролем специального белка - регулятора . Оперон состоит из: 1)Структурных генов 1-го, 2-х, 3-х и более, которые ответственны за синтез белков и эти белки осуществляют последовательно этапы синтеза какого-либо метаболита - продукт обмена синтеза . 2) Оперон содержит функциональные гены : а)Проматор (П) ген - это участок начала транскрипции. Он определяет какая из целей двойной спирали ДНК будет служить матрицей для синтеза и-РНК. б)Ген-Оператор - это участок ДНК с которым происходит связывание регуляторного белка. в)Ген-Терминатор (Т) - участок в конце оперона, который сигнализирует о прекращении транскрипции.
Ген-Оператор регулирует работу структурных генов, а работу всего оперона контролирует Ген-Регулятор . Ген-Регулятор расположен на участке ДНК (молекулы) и кодирует синтез регуляторского белка, который может быть в состоянии белка-Активатора и белка-Репрессора . Регуляторный белок находится в свободном состоянии не действует на ген-Оператор и оперон работает. Белки синтезируются под контролем структурных генов. По мере накопления метаболитов в клетке они (метаболиты) соединяются с регуляторным белком, изменяют его, превращая в белок репрессор, он связывается с геном-оператором и блокирует оперон, т.е. неработают структурные гены. Синтез белков прекращается и оперон не работает. Для начала транкрипции необходимо присоединие к промотору белка-Активотора, который активизирует работу самого оперона.
52. Понятие и структурных генах, гене регуляторе и операторе и их действия.
Серебровский и Дубинин - доказали сложность структуры гена. Ген - участок молекулы ДНК, в котором заложена информация о структуре белка. Ген является единичец наследственной информации. Функция гена : программирование синтеза белков в клетке, но сам ген непосредственно участие в синтезе белка не принимает, т.е. через белки осуществляется контроль за развитие признака. Ген à Белки à Признаки . У низших организмов один ген определяет развитие одного белка, этот белок определяет развитие одного признака. У высших много генов. Один ген определяет развитие многих генов, а эти белки только один признак.
Гены разделяются на 2-е категории : 1)Структурные гены. 2)Функциональные гены. Структурные гены определяют последовательность аминокислот в первичной структуре белка. Функциональные гены контролируют работу структуры генов. В 1961 году французские учёные Жакобо и Вано разработали теорию регуляции активности генов (это включения или выключения тех или иных генов в процессе синтеза белков на уровне транскрипции) - оперонная регуляция . Оперон - участок молекулы ДНК, на котором осуществляется трунскрипция молекулы и-РНК под контролем специального белка - регулятора . Оперон состоит из: 1)Структурных генов 1-го, 2-х, 3-х и более, которые ответственны за синтез белков и эти белки осуществляют последовательно этапы синтеза какого-либо метаболита - продукт обмена синтеза . 2) Оперон содержит функциональные гены : а)Проматор (П) ген - это участок начала транскрипции. Он определяет какая из целей двойной спирали ДНК будет служить матрицей для синтеза и-РНК. б)Ген-Оператор - это участок ДНК с которым происходит связывание регуляторного белка. в)Ген-Терминатор (Т) - участок в конце оперона, который сигнализирует о прекращении транскрипции.
Ген-Оператор регулирует работу структурных генов, а работу всего оперона контролирует Ген-Регулятор . Ген-Регулятор расположен на участке ДНК (молекулы) и кодирует синтез регуляторского белка, который может быть в состоянии белка-Активатора и белка-Репрессора . Регуляторный белок находится в свободном состоянии не действует на ген-Оператор и оперон работает. Белки синтезируются под контролем структурных генов. По мере накопления метаболитов в клетке они (метаболиты) соединяются с регуляторным белком, изменяют его, превращая в белок репрессор, он связывается с геном-оператором и блокирует оперон, т.е. неработают структурные гены. Синтез белков прекращается и оперон не работает. Для начала транкрипции необходимо присоединие к промотору белка-Активотора, который активизирует работу самого оперона.
53. Оплодотворение и его генетическое действие. Андрогенез, гиногенез и филогенез.
Гаметогенез - общее название процесса развития половых клеток. У животных гаметогенез бывает: диффузным , когда гаметы развиваются в любой части тела (у кишечнополостных). Локализованным , когда гаметы развиваются в половых железах (гонады ). Процесс развития мужских половых клеток, сперматозоидов называется сперматогенез и происходит в семенниках. Процесс образования женских яйцеклеток называется оогенезом и происходит в яйцеклетке. В половых железах различают 3-и зоны: зона размножения; роста; созревание половых клеток .
В зоне размножения находятся диплоидные первичные половые клетки (сперматогонии и оогонии содержащие 2 n -хромосом (46 хромосом) ). Они делятся путём митоза и число их увеличивается и образуется сперматоциты и ооциты (содержат 2 n -хромосом ).
В зоне роста , где клетки не делятся, а увеличиваются в размерах, накапливая питательные в-ва. Клетки в зоне роста называются сперматоциты 1-го порядка и ооциты 1-го порядка . В этой зоне каждая хромосома удваивается и состоит из 2-х пар хроматид, сперматоциты и ооциты 1-го порядка содержат 46 - удвиенных 2 n - хромосом .
В зоне созревания клетки делятся 2-а раза: Мейозом I и II . При развитии сперматозоидов оба деления равны, т.е. образуются после Мейоза I 2-е клетки, сперматоциты 2-го порядка (содержащие n -удвоенных хросомом ) и каждая клетка делится Мейозом II . Образуется 4-е сперматиды (содержащие n -хромосом (23-удвоенных) ). Эти сперматиды превращаются в сперматозоиды. В зоне созревания женских половых клеток также 2-а деления Мейоза Iи II. При первом делении образуются 2-е неравные по размерам клетки: одна крупная - это ооцит 2-го порядка (содержит n-удвоенных хромосом), другая маленькая - 1-ое направительное тельце (содержит n-удвоенное). Второе деление Мейзо IIтакже неравное образуется крупная яйцеклетка и 2-ое направительное тельце. 1-ое направительное тельце делится и образуется 2-а вторых редукционных (направленных) телец. Следовательно после оогенеза после 2-х делений меоза образуется 4-е клетки: 3-мелкие направительные и 1-а крупная - гаплоидная клетка с большим запасом питательных в-в.
Оплодотворение - это слияние ♀ и ♂ половых клеток приводящая к образованию зиготы, которая даёт начало новому организму. Оплодотворение специфический в видовом отношении процесс, т.е. сперматозоиды 1-го вида организма не оплодотворяют яйцеклетку другого вида. Яйцеклетка вырабатывает белковые в-ва опрелённого состава, которые взаимодействуют с веществами находящиеся на поверхности сперматозоида. Процесс оплодотворения заключается в проникновении одного или нескольких сперматозоидов в яйцеклетку. Нужна большая концентрация сперматозоидов (1000 и более) чтобы разрушить оболочку яйцеклетки.
Основные формы взаимодействия аллельных генов – полное и неполное доминирование, сверхдоминирование и кодоминирование
Полное доминирование (доминантность) – полное преобладание в фенотипе гетерозиготного организма одного аллеля (доминантного) над другим (рецессивным) аллелем одного и того же гена. Рецессивность – угнетение в фенотипе гетерозиготного организма одного аллеля (рецессивного) другим аллелем (доминантным) одного и того же гена. Доминирование может быть полным и неполным. В случае полного доминирования доминантная гомозигота (АА) и гетерозигота (Аа) имеют одинаковый фенотип. Явление полного доминирования наблюдалось в опытах Г. Менделя, где один аллельный ген всегда был доминантным, другой – рецессивным. Поэтому семена гороха по цвету всегда были или желтыми, или зелеными и не имели другого, например, синего цвета. При полном доминировании в скрещивании гетерозигот (Аа х Аа) расщепления за фенотипом было – 3:1, за генотипом – 1:2:1.
По типу полного доминирования у человека наследуются менделирующие признаки (моногенное наследования): ямочки на щеках, способность свертывать язык трубочкой, свойство загибать язык назад, свободная мочка уха, а также много наследственных болезней: полидактилия, полидактилия, миопатия, кистозно-аденоидная эпителиома, ахондроплазия, др.
Неполное доминирование – взаимодействие аллельных генов, при котором у гетерозиготного организма доминантный аллель не проявляет полностью своей доминантности, а рецессивний аллель одного и того же гена – своей рецессивности. При неполном доминировании фенотип гетерозиготы Аа есть промежуточным между фенотипом доминантной АА и рецесивной аа гомозигот. Так, в скрещивании ночной красавицы с красными цветками (АА) и ночной красавицы с белыми цветками (аа), все гибриды (Аа) первого поколения F1 имели розовую окраску цветков. В скрещивании гибридов первого поколения F1 между собою (Аа х Аа) во втором поколении F2 происходит расщепления за фенотипом в соотношении 1:2:1, которое совпадает со соответствующим по генотипому 1АА:2Аа:1аа, но отличается от расщепления по фенотипу при полном доминировании (3:1).
По типу неполного доминирования у человека наследуются цистинурия, пильгерова анемия, талассемия, атаксия Фридрейха, др. В гомозигот за рецессивним геном цистинурии аа в почках образуются цистиновые камни, в гетерозигот Аа наблюдается лишь повышенное содержимое цистина в моче, гомозиготы АА – здоровые.
Сверхдоминирование – взаимодействие аллельных генов, при котором доминантный аллель в гетерозиготном состоянии проявляется в фенотипе сильнее, чем в гомозитнм (Аа >АА). При этом типе имеет место действие летальных генов. У человека, например, укороченные пальцы рук – брахидактилия – аутосомно-доминантный признак. При чем гомозиготы доминантные погибают еще на ранних стадиях эмбриогенеза. Следуем, гетерозиготы являются больными брахидактилией, а гомозиготы доминантные имеют нормальное строение кисти. В результате брака у родителей, страдающих брахидактилией могут быть дети больные этим заболеванием и здоровые в соотношении 2:1.
Кодоминирование – взаимодействие аллельных генов, при котором в фенотипе гетерозиготного организма проявляются оба аллеля одного и того же гена. По типу кодоминирования у человека наследуется четвертая группа крови (генотип ІАІВ). У людей с этой группой в эритроцитах крови одновременно имеются антиген А, который контролируется аллелем ІА и антиген В – продукт экспрессии аллеля ІВ. Аллели ІА и ІВ – кодоминантные.