Ang mga Vertebrates ay pinagsama ng isang karaniwang morphophysiological na organisasyon. Sa lahat ng mga organ system ng mga hayop na ito, maaaring masubaybayan ng isa ang mga tampok ng sunud-sunod na pagbabago na may kaugnayan sa ebolusyonaryong pagbabago ng mga homologous na organo. Nasa ibaba ang isang pangkalahatang plano ng istraktura, paggana at pagtula sa ontogeny ng mga indibidwal na organ system.
Balat kumakatawan sa isang napakahalagang functional system. Ang balat ay direktang nakikipag-ugnayan sa panlabas na kapaligiran at direktang apektado nito. Ang balat at mga kalamnan ay bumubuo sa katawan ng hayop mula sa ibabaw, bigyan ito ng hugis at hawakan ang lahat ng mga panloob na organo. Pinoprotektahan ng mga takip ng balat ang katawan mula sa panlabas na mekanikal at kemikal na pinsala, pagkakalantad sa temperatura, pagkatuyo, pagtagos ng mga mikrobyo. Ang balat ay nakikibahagi sa thermoregulation, gas exchange at excretion ng mga produkto ng pagkabulok. Ang mga derivatives ng balat ay maaaring makilahok sa pagbuo ng mga organo ng lokomotion (hooves), nagsisilbi para sa paghawak (claws), pag-atake at pagtatanggol (mga sungay, karayom, atbp.), Paglipad (folds), paglangoy (webs).
Ang balat ay naglalaman ng mga receptor ng mga organo ng pagpindot, mayroon itong maraming mga glandula para sa iba't ibang layunin (mucous, mataba, mabaho, pawis, atbp.).
Ang balat ng Vertebrate ay nailalarawan sa pamamagitan ng dalawang layer. Ang panlabas na layer - ang epidermis - ay mula sa ectodermal na pinagmulan. Ito ay palaging multilayered. Ang mas mababang layer nito ay nananatiling buhay at aktibo para sa buhay at gumagawa ng mga bagong layer ng mga cell. Ang itaas na mga layer ng epidermis ay karaniwang binubuo ng mga piping selula, na sa mga terrestrial vertebrates ay nagiging keratinized, namamatay at patuloy.
slough off. Ang epidermis ay nagbubunga ng mga malibog na derivatives ng balat - malibog na kaliskis, balahibo, hairline, claws, hooves, guwang na sungay. Ang iba't ibang mga glandula ng balat ay nabubuo sa epidermis.
Ang panloob na fibrous layer ng balat corium, kung hindi man ay tinutukoy bilang cutis, o ang balat mismo, ay bubuo mula sa mesodermal rudiment - ang balat ng dahon ng somite. Ang layer ng corium ay makapal, ito ay bumubuo sa pangunahing bahagi ng balat, na may mahusay na lakas. Sa corium, ang iba't ibang mga ossification ay nabubuo sa anyo ng mga kaliskis ng isda, mga buto ng integumentaryo, na bumubuo ng tinatawag na balangkas ng balat (sa kaibahan sa chondral skeleton). Sa kapinsalaan ng balat mismo, ang mga sungay ng buto ng usa ay bubuo din. Sa ibabang bahagi ng cutis, isang subcutaneous layer ng taba ang naipon.
kalamnan. Ang layer ng mga kalamnan na matatagpuan sa ilalim ng balat ay ang bulk ng kalamnan, na tinatawag na kalamnan ng katawan, o somatic. Nagbibigay ito ng mga hayop na may kakayahang lumipat sa kapaligiran at binubuo ng striated muscle tissue. Sa mas mababang mga vertebrates, tulad ng sa mga hindi cranial, ang kalamnan ay may naka-segment na karakter. Sa mas mataas na vertebrates, dahil sa pangkalahatang komplikasyon ng mga paggalaw ng katawan, na may pag-unlad ng mga limbs, ang segmentasyon ay nabalisa, at ang mga kalamnan ng puno ng kahoy ay pinagsama-sama, na bumubuo ng mga bahagi ng katawan tulad ng katawan, ulo, at mga organo ng paggalaw.
Maliban sa mga kalamnan ng somatic Ang mga Vertebrates ay may bituka na kalamnan at iba pa lamang loob(mga sisidlan, mga channel). Ang kalamnan na ito ay tinatawag na visceral. Binubuo ito ng makinis na tisyu ng kalamnan at nagbibigay, sa partikular, ang paggalaw ng pagkain sa mga bituka, pag-urong ng mga dingding. mga daluyan ng dugo.
Trunk musculature embryonicly arises mula sa panloob na layer ng myotome (tingnan ang pag-unlad ng lancelet), i.e. dorsal mesoderm. Ang visceral musculature ay isang derivative ng lateral plate, i.e. mesoderm ng tiyan.
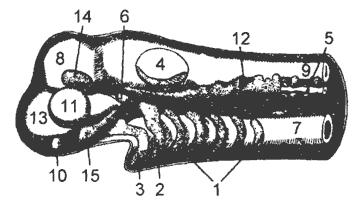
Panloob na balangkas- Ito ang sumusuportang base ng katawan ng isang vertebrate na hayop. Bilang karagdagan, ang balangkas ay kasangkot sa paggalaw ng katawan, pinoprotektahan ang mga panloob na organo.
Sa topograpiya, ang balangkas ng mga vertebrates ay maaaring nahahati sa axial, visceral, limb girdle at libreng mga paa.
Ang axial skeleton sa orihinal nitong anyo ay ipinakita chord napapaligiran ng isang makapal na lamad ng connective tissue. Ang huli ay sumasaklaw hindi lamang sa chord, kundi pati na rin sa neural tube na nakahiga sa itaas nito. Ang notochord ay bubuo mula sa rudiment ng dorsal side ng pangunahing bituka, i.e. ay nagmula sa endodermal.
kanin. 9. Pagbuo ng vertebra (mga nakahalang seksyon):
1 - ang simula ng mas mababang arko; 2 - ang simula ng itaas na arko; 3 - rib, 4 - proseso ng hemal, 5 - lateral na proseso ng vertebra, 6 - superior arch
Sa karamihan ng mga vertebrates, ang notochord ay inilipat at pinalitan ng isang cartilaginous o bony skeleton. cartilaginous at kalansay ng buto s nabubuo bilang mga derivatives ng nasa itaas na connective tissue (mesoderm in origin) membrane. Ang shell na ito ay kaya skeletal.
Sa axial skeleton mayroong vertebral column at cerebral skull. Sa panahon ng pag-unlad ng vertebrae, ang metamerically located paired cartilages na katabi ng ibabaw ng notochord ay unang inilatag. Ito ang mga simulain ng upper at lower vertebral arches (Fig. 9.1). Ang paglaki at pagsasara ng mga panlabas na dulo ng itaas na mga arko ay humahantong sa pagbuo spinal canal, kung saan matatagpuan ang neural tube (Larawan 9, II). Ang mas mababang mga arko ay malapit sa rehiyon ng caudal (sa isda) at limitasyon hemal canal, saan dumadaan ang dorsal aorta at tail vein. Bilang resulta ng pagsasara ng mga panloob na dulo ng itaas at mas mababang mga arko, ang mga vertebral na katawan ay nabuo, sa loob at sa pagitan ng mga ito, sa ilang mga lawak, ang isang notochord ay maaaring mapangalagaan (Larawan 9, III). Sa rehiyon ng puno ng kahoy, ang mga buto-buto ay nakakabit sa mga proseso ng mas mababang vertebral arches.
bungo ng utak, o cranium, ay inilatag sa anyo ng dalawang pares ng cartilage na nakahiga sa ilalim ng rudiment ng utak. Ang kanilang pares sa likod - parachordalia- matatagpuan sa mga gilid ng front end ng chord; pares sa harap - trabeculae- sa harap niya. Binabago nila ang mga simulain ng unang vertebrae at bahagyang ang mga arko ng visceral skeleton. Ang paglaki at pagsasara ng parachordalia at trabeculae ay humahantong sa pagbuo ng pangunahing lamina ng bungo, na sumasailalim sa utak. Kasabay nito, lumilitaw ang mga cartilaginous capsule sa paligid ng mga sensory organ (amoy, paningin, pandinig) na nabuo. Ang mga ito ay matatagpuan nang bahagya sa itaas ng antas ng pangunahing
kanin. 10. Pagbuo ng Shark Cartilaginous Skull:
1 - mga arko ng gill; 2 - hyoid arch; 3 - arko ng panga; 4 - pandinig na kapsula; 5 - chord; 6 - pituitary gland; 7 - bituka; 8 - midbrain; 9 - spinal cord; 10 - butas ng ilong; 11 - mata; 12 - parachordalia; 13 - pangunahing forebrain; 14-orbital cartilages; 15 - trabeculae
mga plato ng bungo at tinatakpan ang utak mula sa mga gilid. Sa kasunod na pag-unlad, ang mga kapsula ng mga organo ng pandama ay konektado at pinagsama sa pangunahing lamina ng bungo. Sa cartilaginous na estado ng bungo, ang isang buong bubong ay hindi nangyayari sa braincase. Ang mga butas na natitira sa pagitan ng mga lintel ng bubong ng bungo - fontanelles- Hinihigpitan ng isang lamad ng nag-uugnay na tissue. Ang isang solidong bubong ng bungo ay nangyayari lamang na may kaugnayan sa pagbuo ng mga superimposed (balat) na buto (frontal, parietal).
Kaya, ang bungo ng utak ay lumitaw na may kaugnayan sa pag-unlad ng utak at pandama na mga organo bilang kanilang proteksiyon na pormasyon.
Visceral skeleton phylogenetically nabuo nang nakapag-iisa sa cranium (Fig. 10). Ang pagtula nito ay nangyayari sa connective tissue malapit sa harap ng digestive tube. Sa una, ang visceral skeleton ay isang serye ng maraming monotonous arches na matatagpuan sa pagitan ng gill slits. Nagsisilbi silang suporta para sa respiratory apparatus.
Ang kasunod na pagbabagong-anyo ng visceral skeleton ay nauugnay sa pagkuha ng mga vertebrates ng mga organo tulad ng upper at lower primary jaws, gitnang tainga, sahig ng bungo ng utak, at larynx.
Ang konsepto ng "visceral skeleton" ay pangunahing itinuturing na may kaugnayan sa mas mababang mga vertebrates. Sa mas mataas na edukasyon ito ay pinalitan ng mga konsepto ng "visceral skull", "facial skull".
Skeleton ng limb girdle at free limbs. Sa mga vertebrates, nakikilala ang hindi magkapares at magkapares na mga paa. Sa turn, ang magkapares na mga limbs ay maaaring alinman sa mga palikpik o ground-type na mga paa.
Skeleton ng hindi magkapares na mga paa - dorsal, caudal, anal fins- binubuo ng ilang cartilaginous o bone rays na hindi nauugnay sa ibang bahagi ng balangkas.
Ang balangkas ng magkapares na limbs ay nahahati sa balangkas ng mga sinturon ng paa at ang balangkas ng malayang paa. Ang mga sinturon ng paa ay laging matatagpuan sa loob ng katawan ng hayop. Ang free limb skeleton sa mga vertebrates ay may dalawang uri: palikpik ng isda at

kanin. 11. Scheme ng vertebrate skeletons:
I - balangkas ng isda, II - balangkas ng terrestrial vertebrate
limang daliri sa paa terrestrial vertebrates. Sa unang kaso, ang balangkas ay kinakatawan ng ilang mga hilera ng kartilago o mga buto na gumagalaw na may kaugnayan sa sinturon bilang isang solong pingga. Ang balangkas ng isang limang-daliri na paa ay binubuo ng isang bilang ng mga lever na maaaring ilipat ang parehong magkasama na may kaugnayan sa sinturon ng paa, at hiwalay - isang kamag-anak sa isa (Fig. 11). Ang pagtula ng balangkas ng paa ay nangyayari sa connective tissue layer ng balat.
Mga organong pantunaw. Ang sistema ng pagtunaw ay kinakatawan ng isang tubo na nagsisimula sa bibig at nagtatapos sa anus. Ang digestive tract ay nabuo mula sa endodermal tube ng gastrula (tingnan ang pag-unlad ng lancelet). Sa bagay na ito, ang epithelium ng digestive tract ay endodermal. Tanging sa rehiyon ng oral at anal openings ang endodermal epithelium ay hindi mahahalata na pumasa sa ectodermal. Ito ay dahil sa invagination ng mga dingding ng katawan (at, dahil dito, ang ectoderm) sa panahon ng pagbuo ng mga butas sa itaas.
Ang digestive tract ay nahahati sa mga sumusunod na pangunahing seksyon: 1) oral cavity paghahatid para sa pagkain; 2) lalaugan- isang departamento na palaging nauugnay sa mga organ ng paghinga: sa isda, ang mga gill slits ay nakabukas sa pharynx, sa mga terrestrial vertebrates, ang isang laryngeal slit ay matatagpuan sa pharynx; ang pharynx ay wastong tinatawag na seksyon ng paghinga ng tubo ng pagtunaw; 3) esophagus, 4) tiyan- pagpapalawak ng bituka ng bituka, na sa ilang mga kaso ay may isang napaka-komplikadong aparato; 5) bituka, karaniwang nahahati sa anterior, o manipis, gitna, o makapal, at hulihan, o tumbong. Ang morphological complication ng intestinal tract sa isang serye ng mga vertebrates ay sumusunod sa landas
ang pagpahaba at pagkakaiba nito sa mga departamento. Ang mga duct ng tatlong uri ng digestive gland ay bumubukas sa digestive tube: laway, atay, pancreas.
Mga glandula ng laway- Pagkuha ng mga terrestrial vertebrates. Binabago nila ang mga mucous glands ng oral cavity. Ang kanilang sikreto ay nagbabasa ng pagkain at nagtataguyod ng pagkasira ng mga carbohydrate.
Ang atay at pancreas ay nabubuo sa pamamagitan ng pag-usli ng anterior na bahagi ng embryonic gut. Ang atay ay nagmumula sa isang bulag na paglaki ng dingding ng tiyan ng bituka (tingnan ang hepatic outgrow ng bituka ng lancelet). Ang mga duct ng atay ay umaagos sa nauuna na maliit na bituka. Pancreas bubuo mula sa ilan, mas madalas mula sa tatlo, mga simulain, na mga paglaki rin ng bituka. Ang glandula na ito, hindi katulad ng atay, ay karaniwang walang hitsura ng isang compact na katawan, at ang mga lobules nito ay nakakalat sa buong mesentery ng anterior maliit na bituka.
Ang mga function ng parehong mga glandula ay mas malawak kaysa sa digestive. Kaya, ang atay, bilang karagdagan sa pagtatago ng apdo, pag-emulsify ng mga taba at pag-activate ng pagkilos ng iba pang mga digestive enzymes, ay isang mahalagang metabolic organ. Dito, ang ilang mga nakakapinsalang produkto ng pagkabulok ay neutralisado, ang glycogen ay naipon. Sinisira ng mga pancreatic enzyme ang mga protina, taba at carbohydrates. Kasabay nito, ang pancreas ay nagsisilbing isang organ panloob na pagtatago. Ang pagkasira sa function na ito ay humahantong sa pagkawala ng kakayahan ng katawan na gumamit ng asukal. Ang resulta ay isang malubhang sakit - diabetes.
Sistema ng paghinga Mayroong dalawang uri ng vertebrates - hasang at baga, at sa isang makabuluhang bahagi ng vertebrates, ang balat ay mahalaga sa paghinga.
Ang gill apparatus ay isang sistema ng magkapares, kadalasang simetriko ang kinalalagyan, slits na nagsisilbing komunikasyon ng pharynx sa panlabas na kapaligiran. Ang anterior at posterior wall ng gill slits ay may linya na may mucous membrane na bumubuo ng lamellar outgrowths; ang mga bunga ay nahahati sa petals, dala ang pangalan hasang. Ang bawat gill plate sa itaas ng mga petals ay pinangalanan kalahating hasang. Sa pagitan ng mga gill slits (sa gill septa) ay nabubulok visceral gill arches(Tingnan ang p. 27 para sa visceral skeleton). Kaya, ang bawat gill arch ay konektado sa dalawang kalahating hasang ng dalawang magkaibang hasang slits.
Gill slits ay inilatag sa anyo ng isang sistema ng mga ipinares na endodermal protrusions na lumalaki palabas mula sa pharynx. Kasabay nito, lumilitaw ang ectodermal invaginations ng panlabas na integument. Ang primordia ay lumalaki patungo sa isa't isa at pagkatapos ay magkaisa. Samakatuwid, ang gill slits ay may pinaghalong ento- at ecto-dermal na pinagmulan. Gill filament Karaniwan
nabubuo mula sa ectodermal rudiment ng fissure, at sa mga hayop na walang panga lamang sila ay may pinagmulang endodermal.
Mga organo ng paghinga ng terrestrial vertebrates - baga- sa scheme sila ay isang pares ng mga bag na nagbubukas sa pharynx sa pamamagitan ng laryngeal fissure. Embryonally, ang mga baga ay bumangon sa anyo ng isang protrusion ng dingding ng tiyan ng pharynx sa likod ng gill apparatus, i.e. ay nagmula sa endodermal. Sa mga unang yugto ng pag-unlad ng embryonic, ang mga buds ng baga ay kahawig ng isang pares ng panloob (endodermal) gill slits. Ang mga pangyayaring ito, pati na ang suplay ng dugo at mga tampok ng innervation na karaniwan sa mga baga at hasang, ay nagpipilit sa amin na isaalang-alang ang mga baga bilang mga homologue ng posterior na pares ng mga gill sac.
Balat nakikilahok sa paghinga sa mga kaso kung saan walang siksik na sungay o bony na kaliskis dito, halimbawa, sa mga amphibian, hubad na balat na isda.
Sa paggana, ang sistema ng paghinga ay kasangkot sa pagpapayaman ng dugo na may oxygen at sa pag-alis ng carbon dioxide. Ang ammonia ay inilalabas sa pamamagitan ng respiratory system sa mas mababang mga hayop sa tubig. Sa mga hayop na may mainit na dugo, ito ay kasangkot sa mga proseso ng thermoregulation. Prinsipyo ng operasyon sistema ng paghinga-pagpapalit ng CO 2 at O 2 sa pagitan ng mga daloy ng gas at dugo na nakadirekta sa countercurrent patungo sa isa't isa.

kanin. 12. Circulatory scheme ng isda (I) at terrestrial vertebrate (II):
1 - branchial arteries; 2 - carotid artery; 3 - dorsal aorta; 4 - aorta ng tiyan; 5 - pulmonary artery; 6 - Cuvier duct; 7 - anterior cardinal (jugular) vein; 8 - posterior cardinal vein; 9 - posterior vena cava; 10 - hepatic vein; 11 - pulmonary vein; 12 - portal vein ng atay; 13 - subintestinal vein
Mga organo ng sirkulasyon. Ang sistema ng sirkulasyon sa mga vertebrates, tulad ng sa mga hindi cranial, ay sarado. Ang sistema ng sirkulasyon ay inilatag mula sa mga panloob na sheet ng mga lateral plate (tingnan ang pag-unlad ng lancelet). Binubuo ito ng magkakaugnay na mga daluyan ng dugo, na sa isang magaspang na pamamaraan ay maaaring mabawasan sa dalawang putot: likod kung saan dumadaloy ang dugo mula ulo hanggang buntot, at tiyan kung saan ito gumagalaw sa kabilang direksyon. Hindi tulad ng non-cranial (Fig. 12), sa mga vertebrates, ang paggalaw ng dugo ay nauugnay sa aktibidad ng puso.
Ang puso ay isang makapal na pader na muscular bag, nahahati sa ilang mga seksyon - mga silid. Ang mga pangunahing bahagi ng puso ay ang atrium, na tumatanggap ng dugo, at ang ventricle, na nagdidirekta nito sa katawan. Ang bilang ng mga silid ng puso ay iba sa iba't ibang klase ng mga vertebrates.
Sa embryonally, ang puso ay bumangon bilang isang pagpapalawak ng posterior na bahagi ng aorta ng tiyan, na sa lugar na ito ay pumulupot sa isang hubog na loop. Ang nauunang bahagi ng loop ay nagbibigay ng pagtaas ventricle puso, likod - atrium.
Ang puso ay may mga striated na kalamnan na gumagana sa awtomatikong mode, at ang pag-urong nito ay hindi napapailalim sa mga volitional impulses. Ang mga sukat ng puso ay nauugnay sa intensity ng trabaho nito, at ang mga sukat nito na nauugnay sa mga sukat ng pagtaas ng katawan sa serye ng mga vertebrates (Talahanayan 1).
Talahanayan 1
Cardiac index sa mga vertebrates ng iba't ibang klase
Ang mga daluyan ng dugo ay nahahati sa dalawang sistema: arterial kung saan dumadaloy ang dugo mula sa puso, at kulang sa hangin kung saan ang dugo ay bumalik sa puso. Sa proseso ng komplikasyon ng mga vertebrates, mayroong isang paglipat mula sa mga hayop na may isang bilog ng sirkulasyon ng dugo sa mga may-ari ng dalawang bilog ng sirkulasyon ng dugo.
Sa likas na katangian nito, ang dugo ay tumutukoy sa nag-uugnay na tisyu na tumagos sa daluyan ng dugo mula sa intercellular space.
Ang dugo ng mga vertebrates ay binubuo ng isang walang kulay na likido - plasma, kung saan matatagpuan ang mga nabuong elemento ng dugo: mga pulang selula ng dugo, o mga erythrocytes, na naglalaman ng isang tina - hemoglobin, at mga puting selula ng dugo - mga leukocytes. Ang mga pulang selula ng dugo ay nauugnay sa oksihenasyon ng dugo, dahil nagdadala sila ng oxygen. Ang mga leukocyte ay kasangkot sa pagkasira ng mga microorganism na pumasok sa katawan. Bilang karagdagan, may mga platelet sa dugo, na may mahalagang papel sa pamumuo ng dugo, pati na rin ang iba pang mga selula. Ang masa ng dugo sa ebolusyonaryong serye ng mga vertebrates ay tumataas (Talahanayan 2).
talahanayan 2
Relatibong masa ng dugo sa masa ng katawan sa mga vertebrates ng iba't ibang klase
(pagkatapos ng Prosser at Brown, 1967; pagkatapos ng Prosser, 1978)
Ang sistema ng sirkulasyon ay multifunctional. Ito ay kasangkot sa pagtanggap ng mga organo, tisyu, mga selula ng oxygen, organic at mineral, mga likido at ang pag-alis ng mga produkto ng pagkabulok, lason, carbon dioxide, sa paglipat ng mga hormone ng mga glandula ng endocrine, atbp.
Kasama ng circulatory system, ang mga vertebrates ay may isa pang vascular system na nauugnay dito - lymphatic. Binubuo ito ng lymphatic vessels at lymph glands. Ang lymphatic system ay hindi sarado. Tanging ang malalaking sisidlan nito ay may independiyenteng mga pader, habang ang kanilang mga sanga ay bumubukas sa mga intercellular space ng iba't ibang organo. Ang mga lymphatic vessel ay naglalaman ng walang kulay na likido - lymph, kung saan ang mga lymphocyte, na nabuo sa mga lymph glandula, ay lumulutang. Ang paggalaw ng lymph ay natutukoy sa pamamagitan ng pag-urong ng mga pader, ilang mga seksyon ng malalaking sisidlan (ang tinatawag na lymphatic hearts) at pana-panahong pagbabago ng presyon sa mga sisidlan ng iba't ibang organo.
Ang lymphatic system ay nagsisilbing isang tagapamagitan sa pagpapalitan ng mga sangkap sa pagitan ng dugo at mga tisyu.
Sistema ng nerbiyos. Ang mga pag-andar ng sistema ng nerbiyos ay ang pang-unawa ng panlabas na stimuli at ang paghahatid ng mga umuusbong na paggulo sa mga cell, organo, tisyu, pati na rin ang pag-iisa at koordinasyon ng mga aktibidad ng mga indibidwal na organ system at ang katawan sa kabuuan sa isang solong gumaganang pamumuhay. sistema. Embryonally sistema ng nerbiyos sa mga vertebrates, ito ay bumangon, tulad ng sa mga hindi cranial, sa anyo ng isang guwang na tubo na inilatag sa ectoderm sa dorsal na bahagi ng embryo (Larawan 13). Kasunod nito, nangyayari ang pagkakaiba-iba nito, na humahantong sa pagbuo ng: a) central nervous system, kinakatawan ng utak at spinal cord, b) peripheral nervous system, na binubuo ng mga nerbiyos na umaabot mula sa utak at spinal cord, at c) sympathetic nervous system, na binubuo sa batayan ng mga nerve node na matatagpuan malapit sa spinal column at konektado ng mga longitudinal strands.
Ang utak ay kinakatawan sa mga vertebrates ng limang mga seksyon: anterior, intermediate, middle, cerebellum at oblong utak. Ito ay inilatag embryonic sa anyo ng pamamaga ng anterior na bahagi ng neural tube, na sa lalong madaling panahon ay nahahati sa tatlong pangunahing cerebral vesicle (Fig. 14). Sa hinaharap, ang unang cerebral bladder ay nagbubunga sa harap ng forebrain; ang likod na bahagi nito ay binago sa diencephalon. Ang midbrain ay nabuo mula sa pangalawang cerebral bladder. Sa pamamagitan ng protrusion
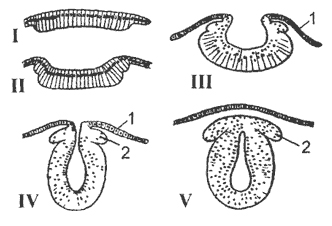
kanin. 13. Mga sunud-sunod na yugto ng pag-unlad ng central nervous system ng mga vertebrates (schematic cross sections):
I-II pagkita ng kaibhan ng ectoderm; III invagination ng neural plate, IV-V - paghihiwalay ng neural tube, 1 epidermis; 2 ganglion plate
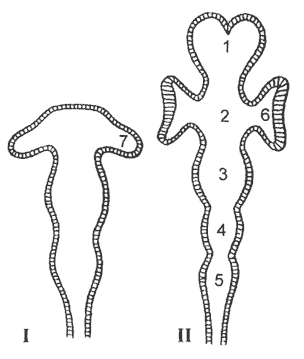
kanin. 14. Pag-unlad ng mga bahagi ng utak (scheme):
I - yugto ng tatlong bula (na may mga bula sa mata), II - yugto ng limang departamento (na may mga tasa ng mata), 1 - forebrain, 2 - diencephalon; 3 - midbrain; 4 - cerebellum; 5 - medulla oblongata; 6 - tasa ng mata; 7 - bula ng mata
ang bubong ng ikatlong cerebral bladder ay bumubuo ng cerebellum, kung saan matatagpuan ang medulla oblongata. Ang forebrain ay higit pang nahahati sa kaliwa at kanang kalahati.
Kasabay ng paglaki at pagkakaiba-iba ng seksyon ng ulo ng neural tube, nangyayari ang isang kaukulang pagbabago ng neurocoel. Ang dalawang extension nito sa forebrain hemispheres ay kilala bilang lateral ventricles utak. Ang pinalawak na bahagi ng neurocoel sa intermediate na bahagi ng utak ay itinalaga bilang ikatlong ventricle, lukab ng midbrain - bilang sylvian aqueduct, ang lukab ng medulla oblongata - bilang ikaapat na ventricle, o rhomboid fossa(Larawan 14). 10 o 12 pares ang umaalis sa utak cranial nerves.
Ang forebrain ay may dalawang simetriko na matatagpuan na protrusions sa harap, kung saan ang unang pares ng mga nerbiyos sa ulo ay umaalis - olpaktoryo(ako). Mula sa ilalim ng diencephalon umalis biswal nerbiyos (pangalawang pares ng mga nerbiyos sa ulo, II).
Sa bubong ng diencephalon, dalawang protrusions na nakaupo sa mga binti ay nabuo: anterior - parietal organ at likod - epiphysis.
Ang isang hindi magkapares na protrusion ay lumalaki mula sa ilalim ng diencephalon - funnel, na katabi ng isang pormasyon na kumplikado sa istraktura at pag-andar - pituitary. Ang anterior pituitary gland ay bubuo mula sa epithelium ng oral cavity, ang posterior - mula sa medulla. Ito ay matatagpuan doon hypothalamus.
Ang bubong ng midbrain ay bumubuo ng mga ipinares na pamamaga - mga visual na lobe (tubercles). Ang ikatlong pares ng mga nerbiyos sa utak ay umaalis sa midbrain ( oculomotor, III). Ikaapat na pares ng mga ugat sa ulo ( harangan, IV) ay umaalis sa hangganan sa pagitan ng gitna at medulla oblongata, ang lahat ng iba pang mga nerbiyos sa ulo (V - X - XII) ay umaalis mula sa medulla oblongata.
Spinal cord hindi nalilimitahan nang husto mula sa medulla oblongata. Sa gitna ng spinal cord (kasama ang pangunahing axis ng organ), ang isang neurocoel ay napanatili, na kilala sa mga vertebrates sa ilalim ng pangalan spinal canal.
Ang mga nerbiyos ng gulugod ay umaalis mula sa spinal cord sa metamerically (ayon sa bilang ng mga segment). Nagsisimula sila sa dalawang ugat: dorsal - pandama at tiyan - motor. Ang mga ugat na ito ay nagsasama sa ilang sandali pagkatapos lumabas sa spinal cord upang mabuo ang mga nerbiyos ng gulugod, na pagkatapos ay nahahati muli sa mga sanga ng dorsal at tiyan.
mga organo ng pandama. Ang pangkat ng mga organo na ito ay lumitaw bilang mga derivatives ng iba't ibang bahagi ng embryo at sa iba't ibang yugto pag-unlad nito. Ito ang mga organo ng pang-amoy, paningin, pandinig, vestibular apparatus, lateral line organs, organo ng panlasa, pagpindot, mga partikular na organo na nakikita ang magnetic field ng Earth, mga electric field, thermal radiation atbp.
Mga organo ng olpaktoryo. Ito ay pinaniniwalaan na ang pang-amoy ay isa sa mga pinaka sinaunang pag-andar ng utak. Ang mga organo ng amoy ay inilalagay bilang isang pampalapot ng ectoderm nang sabay-sabay sa neural plate. Sa kahanay, ang balangkas ng mga kapsula ng olpaktoryo, na bahagi ng bungo ng utak, ay nabuo. Sa una, ang mga olfactory capsule ay nakikipag-usap lamang sa panlabas na kapaligiran at may mga panlabas na butas ng ilong. Kasunod nito, dahil sa pagkakaroon ng terrestrial, ang mga butas ng ilong ay dumaan.
mga organo ng paningin nabibilang din sa mga sinaunang sensory organ. Ang light-sensitive na pagtanggap ay nangyayari sa napakaagang yugto sa ebolusyon ng mga chordates (tandaan ang lancelet) at nabuo sa maagang embryogenesis. Ang mga organo ng paningin ng mga vertebrates ay nahahati sa magkapares at hindi magkapares. Parehong outgrowth ng diencephalon. Ang mga ipinares na mata ay inilalagay bilang mga outgrowth ng mga lateral na bahagi ng diencephalon, hindi magkapares - bilang sunud-sunod na matatagpuan sa bubong ng diencephalon (pineal gland at parietal organ). Ang pagtula ng mga ipinares na mata ay sinamahan ng pagbuo ng mga visual na kapsula sa kanilang paligid, na bahagi ng bungo ng utak (Larawan 15, 16).
mga organ ng pandinig may kumplikadong pinagmulan sa mga vertebrates. Ang pinakamaagang sa ebolusyon ay ang panloob na tainga, na

kanin. 15. Mga sunud-sunod na yugto ng pag-unlad ng mata:
1 - diencephalon; 2 - ang epidermis na bumubuo sa lens; 3 - ang simula ng pagbuo ng tasa ng mata; 4 - binti ng tasa ng mata; 5 - lens; 6 - kornea; 7 - retina; 8 - pigment shell; 9 - mesoderm cells na bumubuo sa choroid at sclera
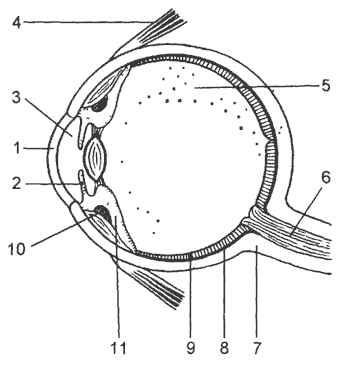
kanin. 16. Sagittal na seksyon ng mata ng tao:
1 - kornea; 2 - iris; 3 - nauuna na silid; 4 - isa sa mga kalamnan na gumagalaw sa mata; 5 - vitreous body; 6 - optic nerve; 7 - sclera; 8 - choroid; 9 - retina; 10 - kalamnan ng ciliary; 11 - ciliary body
ay inilatag sa ectoderm ng embryo, lumalalim sa anyo ng isang butas at nagiging hugis bilang pandinig na vesicle nakahiga sa auditory capsule. Ang auditory vesicle ay nahahati sa pamamagitan ng constriction sa dalawang bahagi. Itaas na seksyon nagiging vestibular apparatus. Ito ang organ ng balanse. Pinapayagan ka nitong madama ang posisyon ng katawan sa tatlong-dimensional na espasyo ng Earth. Ang katawan na ito ay 3 kalahating bilog na kanal sa panloob na tainga (Larawan 17, 18). Ang mas mababang bahagi ng auditory vesicle ay ang panloob na tainga mismo - auditory sac.
Ang gitna at panlabas na tainga ay nabuo sa mga huling yugto ng paglitaw ng mga vertebrates na may kaugnayan sa landfall.
Mga organo ng lateral line katangian lamang ng pangunahing aquatic vertebrates, ay inilatag din sa ectoderm. Ang mga ito ay mga uka na umaabot

kanin. 17. Mga sunud-sunod na yugto ng pag-unlad ng panloob na tainga:
I - auditory placode, II - fossa, III at IV - vesicle sa seksyon; V at VI - ang pagbuo ng mga kalahating bilog na kanal; 1 - simula ng isang snail; 2 - kalahating bilog na kanal; 3 - endolymphatic duct; 4 - bilog na bag; 5 - hugis-itlog na supot
sa gilid ng ulo at sa kahabaan ng katawan. Ang mga uka na ito ay maaaring sakop o hindi ng mga buto-buto. Nakikita ng mga lateral line organ ang magaan na paggalaw at mga panginginig ng tubig na malapit sa mga pinagmumulan ng mga panginginig ng boses: ang bilis at direksyon ng mga alon, mga paggalaw ng sariling katawan at ang pagkakaroon ng mga bagay sa landas ng hayop sa tubig. Ang mga ito ay isang seismic sensor system.
mga organo ng panlasa ay inilalagay sa endoderm at nakikita ang lasa ng pagkain na natupok sa hanay: matamis, mapait, maasim, maalat. Matatagpuan ang mga ito sa mga taste buds sa loob ng oral cavity.
mga organo ng pandama. Wala silang mga sensory cell, ngunit ang mga nerve ending ay sumasanga sa balat, na nakikita ang mga bagay sa kapaligiran sa pamamagitan ng pagpindot.
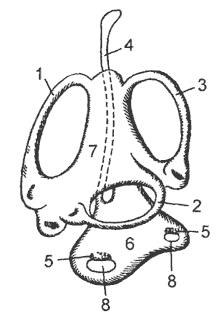
kanin. 18. Scheme ng membranous auditory labyrinth:
1, 2, 3 - anterior, external at posterior semicircular canals, 4 - endolymphatic duct, 5 - auditory spot, 6 - round sac, 7 oval sac, 8 - otoliths

kanin. 19. Bowman's capsule na may vascular tangle:
1 - afferent blood vessel, 2 - efferent blood vessel, 3 - Bowman's capsule, 4 - vascular tangle, 5 - pangunahing ihi, 6 - renal tubule, 7 - funnel ng renal tubule (nephrostome) na bumubukas sa cavity ng katawan

kanin. 20. Scheme ng urinary system ng vertebrates:
I (lalaki) at II (babae) - mga pating at amphibian, III (lalaki) at IV (babae) - mga reptilya at ibon, V (lalaki) at VI (babae) - mga mammal, 1 - pronephros (pronephros); 2 - testis ; 3 - vas deferens, 4 - mesonephros (pangunahing bato), 5 - hindgut, 6 - pantog, 7 - cloaca, 8 - funnel, 9 - ovary, 10 - Muller canal, 11 - epididymis (natitira sa anterior na bahagi ng meso - nephros), 12 - vas deferens, 13 - metanephros (pangalawang bato), 14 pangalawang ureter, 15 mesonephros rudiment, 16 oviduct, 17 ovum, 18 protina na itinago ng mga glandula ng oviduct wall, 19 - uterus, 20 - embryo sa matris 21 - puki, 22 - genital sinus, 23 - prostate glands; 24 - seminal vesicle, 25 - vas deferens, 26 - epididymis, 27 - perineum; 28 - rudiment ng Muller canal, 29 - canal ng mesonephros (pangunahing bato); 30 - copulatory organ (penis), 31 - anus
excretory organs. Lahat ng vertebrates ay may excretory organs bato idinisenyo upang alisin ang labis na tubig, mga mineral na asing-gamot at mga produkto ng agnas ng metabolismo ng nitrogen sa anyo ng urea o uric acid mula sa katawan. Ang mga ito ay mula sa mesodermal na pinagmulan, na nakalagay sa panlabas na dingding ng mga somite. Gayunpaman, ang istraktura at mekanismo ng paggana ng mga bato sa iba't ibang grupo ng mga vertebrates ay hindi pareho. Sa proseso ng ebolusyon ng mga vertebrates, tatlong uri ng bato ang nagbabago: ulo, o pronephros (pronephros), puno ng kahoy, o pangunahin, bato (mesonephros) At pelvic, o pangalawa, bato (metanephros). Ang iba't ibang uri ng bato ay may iba't ibang prinsipyo ng proseso ng paglabas: paglabas mula sa lukab ng katawan, halo-halong paglabas (mula sa lukab ng katawan at mula sa dugo), at sa wakas, mula lamang sa dugo. Kasabay nito, ang mga pagbabago ay nangyayari sa mekanismo ng reabsorption ng tubig. Ang paglabas ng tubig at mga produkto ng metabolismo ng protina na natunaw dito mula sa lukab ng katawan ay posible dahil sa maraming mga nephrostoma na hugis funnel na bumubukas sa lukab ng katawan. Ang paglabas mula sa dugo ay nangyayari sa pamamagitan ng mga Malpighian na katawan ng mga bato. Ang mga excretory ducts mula sa mga bato ay tinatawag mga channel ng lobo, sila ay pinalitan ureters. Karamihan sa mga vertebrates ay mayroon pantog. Sa pangunahing aquatics, ang ammonia ay maaaring itapon sa pamamagitan ng mga hasang.
Mga organo ng kasarian. Ang mga glandula ng kasarian ng mga vertebrates - ang mga ovary sa mga babae at ang mga testes sa mga lalaki - ay karaniwang ipinares. Nabuo sila mula sa seksyon ng mesoderm sa lugar ng paghahati ng rudiment na ito sa somite at lateral plate.
Sa una (sa walang panga) ang mga glandula ng kasarian ay walang mga excretory duct at ang mga produkto ng reproduktibo ay nahulog sa pamamagitan ng mga rupture sa mga dingding ng mga gonad sa lukab ng katawan, mula sa kung saan sila ay pinalabas sa panlabas na kapaligiran sa pamamagitan ng mga espesyal na pores. Kasunod nito, lumitaw ang genital tract, na sa mga lalaki ay nauugnay sa mga excretory organs (Wolf's canal). At sa mga babae, ito ay gumaganap bilang isang oviduct Mueller channel, na nagpapanatili ng koneksyon ng kabuuan sa panlabas na kapaligiran.
Ebolusyon ng mga organ system
"Wala sa biology
may katuturan kung hindi
tulad ng sa liwanag ng ebolusyon"
F.G. Dobzhansky
Mode ng ebolusyon ng gene
cartilaginous na isda
Sa cartilaginous na isda (mga pating, ray), isang ikaapat na karagdagang silid ang bubuo sa puso - ang arterial cone. Sa loob ng kono mayroong ilang mga pares ng mga balbula na pumipigil sa pagbabalik ng dugo sa ventricle. Bilang karagdagan, ang bagong nabuong kalamnan na tisyu ng puso ay nagpapakinis sa matalim na pumipintig na paggalaw ng dugo at ginagawa itong mas pare-pareho. Mula sa arterial cone, ang dugo ay nakadirekta sa aorta ng tiyan at higit pa sa mga hasang. Sa karamihan ng mga cartilaginous na isda, ang mga hasang ay matatagpuan sa apat na cartilaginous gill arches at sa hyoid arch, at naaayon ay mayroong limang pares ng afferent branchial arteries. Sa mga hayop na may pulmonary na uri ng paghinga, ang ilang branchial arteries ay nabawasan (ang pagbawas ay ang kumpletong pagkawala ng isang organ o tissue), at ang ilan ay nabago sa malalaking daluyan ng dugo. Ang pangunahing mga ugat ay dumadaloy sa isang pares ng mga duct ng Cuvier, at pagkatapos ay sa venous sinus, tulad ng sa lancelet. Ang isa pang capillary network ay lilitaw - ang portal system ng mga bato. Ang bilang ng mga pulang selula ng dugo sa mga cartilaginous na isda ay mas malaki kaysa sa mga cyclostomes, na nag-aambag sa supply ng oxygen sa mga tisyu. Sa unang pagkakataon, lumilitaw ang pali - ang organ ng kaligtasan sa sakit at ang lugar ng pagbuo ng mga selula ng dugo.
Sa bony (pinaka-freshwater) na isda, ang arterial cone ay sumailalim sa pagbawas, ngunit ang base ng aorta (aortic bulb) ay lumawak. Ang bilang ng mga erythrocytes ay mas malaki kaysa sa cartilaginous na isda. May katibayan na ang puso ng bony fish ay walang sympathetic innervation, ngunit ibinibigay lamang sa mga sanga ng vagus nerve, na kumokontrol sa mga contraction ng puso.
Lungfish
Sa mga modernong bony fish, ang lungfish ay interesado; sila ay mga kamag-anak ng mga isda na may palikpik na lobe kung saan nagmula ang mga terrestrial vertebrates. Ang morphophysiological feature ng isda na humihinga gamit ang mga hasang at baga ay ang solusyon sa problema ng makatwirang paggamit ng parehong mga organo ng gas exchange sa tubig at hangin. Ang kanilang mga baga ay nabuo sa likod ng pharynx, i.e. sa likod ng hasang. Ang dugo ay pumapasok sa mga baga sa pamamagitan ng pulmonary artery, na nagmumula sa posterior superior branchial artery. Ang oxygenated (oxygen enriched) na dugo ay bumabalik mula sa mga baga patungo sa puso.
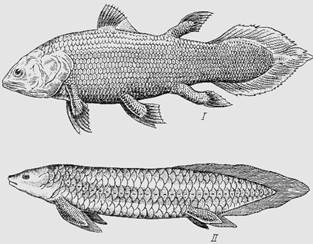
kanin. Loop-finned at lungfish:
I - coelacanth; II - neoceratod
Sa lungfish, ang puso ay binubuo ng isang venous sinus na tumatanggap ng venous blood, isang atrium, isang ventricle, at isang arterial cone. Sa atrium, ventricle at arterial sinus may mga hindi kumpletong panloob na partisyon. Ang venous sinus, na nagdadala ng deoxygenated na dugo mula sa buong katawan, ay dumadaloy sa kanang kalahati ng atrium, at ang mga pulmonary veins, na nagdadala ng dugo na mayaman sa oxygen mula sa mga baga, ay dumadaloy sa kaliwang bahagi ng atrium. Mula sa kaliwang kalahati ng atrium, ang dugo ay pumapasok sa kaliwang kalahati ng ventricle at sa pamamagitan ng arterial cone - sa ibabang bahagi ng pinaikling aorta ng tiyan at pagkatapos ay sa dalawang anterior na pares ng afferent branchial arteries: pagkatapos ay ang dugo ay pumapasok sa ulo sa pamamagitan ng ang mga carotid arteries. Mula sa kanang kalahati ng ventricle, ang deoxygenated na dugo ay nakadirekta sa itaas na bahagi ng abdominal aorta at kasama nito sa dalawang posterior pares ng branchial arteries.
Kaya, ang lungfish ay may pangalawang pulmonary (maliit) na sirkulasyon. Upang maiwasan ang pagpasok ng tubig sa mga baga, ang mga daanan ng hangin ay naharang sa pamamagitan ng pag-urong ng mga kalamnan ng larynx; Napapanahong isara ang puwang na ito mga impulses ng nerve ipinadala ng central rhythm generator sa brainstem. Ang ganitong sistema ng regulasyon ay matatagpuan din sa mga palaka na tadpoles, na humihinga gamit ang mga hasang. Ang mga dayandang ng reaksyong ito ay napanatili sa anyo ng mga hiccups, na kadalasang nangyayari sa mga tao. Ang hiccups ay isang espesyal na paraan ng paghinga kung saan ang biglaang paglanghap ay sinusundan ng mabilis na pagsasara ng glottis. Ang biglaang pagkaputol ng paghinga ay nagdudulot ng tunog na katulad ng "hik". Ang mga pag-atake ng hiccup ay maaaring tumagal ng maraming taon. (Si Charles Osborne ay naobserbahan mula 1922 hanggang 1991, at siya ay nakalista sa Guinness Book of Records).
Dapat itong bigyang-diin na ang hiccups ay isang genetic physiological trait. At napaka sinaunang. Ito ay hindi bababa sa 370 milyong taong gulang. Sa oras na iyon, hindi lamang mga tao, kundi pati na rin ang mga reptilya, ibon at mammal ay wala sa Earth. Kaya, ang mga hiccup ay nag-ugat sa malayong nakaraan at nagpapatotoo sa ating ebolusyonaryong relasyon sa mga isda at amphibian. Ito ay tinatawag na evolutionary inheritance. Hindi lamang mga bata at matatanda ang suminok, kundi pati na rin ang mga embryo na umabot sa edad na dalawang buwan. Ang layunin ng sinaunang reflex na ito sa mga mammalian embryo ay dalawang beses: 1) upang maiwasan ang amniotic fluid mula sa pagpasok sa pagbuo ng mga baga; 2) maghanda para sa pagsuso ng gatas, kung saan kinakailangan ding harangan ang pasukan sa larynx.
Ang mga terrestrial vertebrates ay sumailalim sa mga makabuluhang pagbabago sa istraktura daluyan ng dugo sa katawan, nakabalangkas sa lungfish. Sa lahat ng mga tetrapod, ang aorta ng tiyan (na bahagyang pinalitan ng puso) ay nahahati sa ilang magkakahiwalay na arterial trunks simula sa puso. Ang paghahati ng aorta ng tiyan ay naganap sa iba't ibang paraan sa mga ninuno ng mga modernong amphibian, reptilya, at mammal.
Mga Amphibian (Amphibians)
Ang sistema ng sirkulasyon ng mga amphibian ay unti-unting nagbago kumpara sa parehong sistema ng isda, na may kaugnayan sa pag-landfall at ang pagpapalit ng paghinga ng hasang sa paghinga ng baga. Sa modernong amphibian, tatlong pares ng mga vessel ay nagsisimula mula sa arterial cone ng puso: ang carotid arteries, ang aortic arches (na nagsasama sa dorsal side upang mabuo ang dorsal aorta), at ang pulmonary o cutaneous-pulmonary arteries. Sa pamamagitan ng mga carotid arteries ng amphibian, dumadaloy ang dugo sa ulo; kasama ang aortic arches - sa dorsal aorta, at mula dito sa iba't ibang mga organo ng amphibian body; Ang mga arterya ng pulmonary (o skin-pulmonary) ay nagbibigay ng suplay ng dugo sa respiratory system.
Ang puso ng mga amphibian ay binubuo ng 3 pangunahing seksyon (kaliwa at kanang atria, ventricle) at dalawang karagdagang (venous sinus at arterial cone). Sa mga amphibian, halimbawa, sa isang palaka, ang mga tagaytay ng kalamnan ay nakausli sa cavity ng ventricle mula sa mga dingding nito, na naghahati nito sa mga compartment. Ang pagkakaroon ng mga tagaytay ay bahagyang pumipigil sa paghahalo ng oxygenated at deoxygenated na dugo. Sa panahon ng pulmonary respiration, ang dugo na may mababang nilalaman ng oxygen ay unang pumapasok sa arterial cone at idinidirekta sa balat-pulmonary arteries na pinakamalapit sa puso; ang pangalawang bahagi ng halo-halong dugo ay ipinapadala sa mga arko ng aorta, at ang pangatlo (pinaka-oksiheno) na dugo ay pumapasok sa pamamagitan ng mga carotid arteries patungo sa ulo. Kung ang hayop ay nasa ilalim ng tubig at humihinga sa pamamagitan ng balat, kung gayon mas maraming dugo ang nakadirekta sa balat.
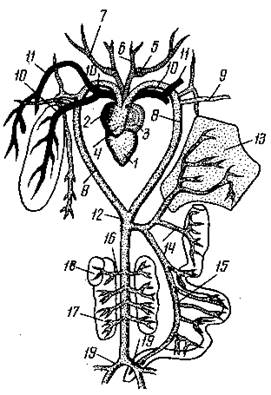
kanin. Scheme ng arterial system ng isang palaka: 1 - ventricle ng puso; 2 - kanang atrium; 3 - kaliwang atrium; 4– arterial cone; 5-7 - mga sanga ng carotid arteries; 8 - mga arko ng mga arterya; 9 - subclavian artery; 10 - pulmonary artery; 11 - isang malaking cutaneous artery; 12 - dorsal aorta; 13 - atay; 14 - gastric artery; 15 - bituka arterya; 16 - arterya ng bato; 17 - bato; 18 - testis; 19 - iliac artery.
(Dapat tandaan na sa isda ang lahat ng mga arko ng gill arteries ay konektado sa bawat isa sa itaas ng pharynx ng mga ugat ng dorsal aorta; ang gayong koneksyon ay napanatili sa ilang mga tetrapod sa anyo ng mga carotid duct sa pagitan ng mga carotid arteries. at ang aortic arches, gayundin ang botallic ducts sa pagitan ng aortic arches at pulmonary arteries. ) Sa mga tao, isang ductus arteriosus lamang ang nag-uugnay sa kaliwang pulmonary artery sa aortic arch sa pagbuo ng fetus. Ang hindi pagsasara ng ductus arteriosus sa isang bagong panganak ay isang atavistic congenital heart disease.
Sa mga aklat-aralin sa paaralan, upang "nakakumbinsi" na ilarawan ang pag-unlad ng sistema mula sa isang simple hanggang sa isang mas kumplikadong organisasyon, ang mga pinasimple na ideya ay ibinibigay tungkol sa istraktura ng puso sa mga isda, amphibian at iba pang mga vertebrates. Ang mga obserbasyon ay nagpapakita na ang istraktura at regulasyon ng cardiovascular system ng mga amphibian ay medyo kumplikado, dahil ang mga hayop na ito ay nakatira sa dalawang kapaligiran - tubig at hangin. Bilang karagdagan, ang ilang mga isda ay may mga stem cell sa kanilang kalamnan sa puso na kasangkot sa myocardial regeneration. Ang mga selulang ito ay wala sa myocardium ng tao, kaya ang tisyu ng peklat ay nabuo sa lugar ng infarction, na pumipigil sa puso na gumana.
mga reptilya
Ang mga reptilya ay may isang pangunahing aromorphosis - ang pagbuo ng isang malakas na pulmonary pump - ang dibdib. Sa pagsasaalang-alang na ito, ang pangangailangan para sa paghinga ng balat ay nawawala at nagiging posible na paghiwalayin ang arterial at venous na daloy ng dugo.
Ang thermoregulation sa mga reptilya ay hindi perpekto, samakatuwid, sila ay nailalarawan sa pamamagitan ng tinatawag na heliothermy - pagpainit ng katawan sa araw. Kapag "sunbathing" kinakailangan upang idirekta ang mas maraming dugo sa balat at mas kaunti sa mga baga, at sa panahon ng masiglang aktibidad - sa mga baga. Alinsunod sa mga pangangailangang ito, ang paggalaw ng dugo sa systemic at pulmonary circulation ay kinokontrol.
Ang aorta ng tiyan sa mga ninuno ng mga reptilya ay nahahati sa tatlong arterial trunks, simula sa puso: ang kaliwa at kanang aortic arches, na pinagsama sa dorsal aorta at ang karaniwang trunk ng pulmonary arteries. Ang kanang arko ng aorta ay tumatanggap ng purong arterial na dugo mula sa ventricle ng puso; sa bagay na ito, ang carotid at subclavian arteries, na nagdadala ng dugo sa ulo at forelimbs, sa mga reptilya ay nagsisimula mula sa kanang arko ng aorta. Ang istraktura ng venous na bahagi ng sistema ng sirkulasyon ay hindi sumailalim makabuluhang pagbabago kumpara sa mga amphibian.
Ang puso ng karamihan sa mga reptilya (mga butiki, ahas, pagong) ay may tatlong silid at binubuo ng 2 ganap na magkahiwalay na atria at isang ventricle. Ang mga pulmonary veins, na nagdadala ng oxygenated na dugo, ay walang laman sa kaliwang atrium. Tatlong vena cava sa kanang atrium. Sa ventricle ng mga reptilya (maliban sa mga buwaya) mayroong isang hindi kumpletong septum. Ang septum ay hindi matatagpuan sa isang patayo (bilang interatrial), ngunit sa isang pahalang na eroplano, at samakatuwid ay naghahati sa ventricle sa mga seksyon ng dorsal at tiyan. Ang parehong atria ay bumubukas sa dorsal ventricle. Dapat pansinin na ang atria sa mga reptilya (hindi katulad ng mga ibon at mammal) ay hindi magkakasabay (halili). Ang kanang atrium ay unang nagkontrata, at ang venous na dugo ay pumapasok sa ventricle ng tiyan, at pagkatapos ay napupunta sa mga baga upang pagyamanin ng oxygen. Ang kaliwang atrium pagkatapos ay kumukuha at pinupuno ng dugo ang dorsal ventricle. Mula sa seksyong ito, nagsisimula ang kanan at kaliwang arko ng aorta, na nagsalubong. Sa panahon ng ventricular systole, ang septum ay ganap na naghihiwalay sa rehiyon ng tiyan mula sa rehiyon ng dorsal, upang ang arterial na dugo ay pumasok sa parehong aortic arches, at ang venous blood ay pumapasok sa pulmonary arteries.
Sa umaga, bumababa ang temperatura ng katawan ng mga reptilya, at ang mga hayop ay napipilitang lumipat sa insolation mode - pagpainit sa araw. (Dapat tandaan na ang curve ng temperatura sa mga tao ay sumusunod sa parehong ritmo tulad ng sa mga reptilya: bumababa ito sa umaga, at tumataas sa gabi). Sa panahon ng insolation, ang lumen ng karaniwang trunk ng pulmonary arteries ay makitid, at ang bahagi ng dugo mula sa tiyan na bahagi ng ventricle ay ipinadala sa dorsal na bahagi, kung saan ito ay naghahalo. Ang simula ng kaliwang aortic arch ay matatagpuan sa kanan at sa ibaba ng simula ng kanan, samakatuwid ang halo-halong dugo ay pumapasok sa kaliwang aortic arch, at ang oxygenated na dugo ay pumapasok sa kanan. Ang carotid at subclavian arteries ay nagsisimula sa kanang aortic arch, kaya ang ulo at forelimbs ng mga reptile ay tumatanggap lamang ng arterial blood.
Kinakailangang bigyang pansin ang katotohanan na ang pahalang na septum sa puso ng mga reptilya ay hindi isang matagumpay (maladaptive) na neoplasma, dahil sa isang pahalang na kumpletong istrukturang dibisyon ng ventricle, ang dugo ay hindi makapasok sa pulmonary arterial trunk.
Ang tunay na apat na silid na puso ay nabuo nang nakapag-iisa sa tatlong linya ng ebolusyon: sa mga buwaya, ibon, at mammal. Ito ay itinuturing na isa sa mga pinakamalinaw na halimbawa ng parallel evolution.
Ang parallel evolution ay ang independiyenteng hitsura ng magkatulad na katangian sa mga organismo ng iba't ibang taxa (species, genera, pamilya, klase, atbp.). Ang paralelismo ay malinaw na nagpapakita ng isang regular, i.e. hindi random na kalikasan ng ebolusyon. Ang hindi pangkaraniwang bagay na ito ay nagpapahiwatig na ang paglipat sa higit pa mataas na lebel Ang organisasyon ay hindi nangyayari sa isang malaking grupo ng mga organismo, ngunit sa ilan, na umuunlad nang magkatulad. Kasabay nito, ang mga indibidwal na palatandaan na bumubuo sa aromorphosis (komplikasyon ng organisasyon) ay maaaring lumitaw nang sabay-sabay, at kung minsan sa iba't ibang oras. Kaya, ang mga progresibong katangian ay maaaring unti-unting maipon hanggang, sa isang (o ilang) grupo, sila ay mabuo nang magkakasama.
Sa mga buwaya, na kabilang sa mas mataas na mga reptilya, isang makabuluhang muling pagsasaayos ng puso ang naganap - a patayo (sagittal) septum na ganap na naghihiwalay sa kanang ventricle mula sa kaliwa. At mula sa kaliwang ventricle ay nagsisimula ang kanan (nababalot ang puso sa kanan) aortic arch, at mula sa kanan - ang kaliwa (enveloping ang puso sa kaliwa) aortic arch at ang karaniwang trunk ng pulmonary arteries. Sa lugar kung saan tumatawid ang kanan at kaliwang aortic arches, mayroong panizza foramen kung saan posible ang paghahalo ng arterial at venous blood. Ang kaliwang aortic arch sa mga buwaya (pati na rin ang mga ibon, na kanilang mga kamag-anak) ay hindi sumanib sa kanang arko sa dorsal aorta, tulad ng sa iba pang mga reptilya, ngunit nagpapatuloy sa celiac artery, na nagbibigay ng dugo sa mga bituka. Ang dorsal aorta ay nabuo lamang ng kanang arko. Kapag ang isang buwaya ay nasa ilalim ng tubig, ang daloy ng dugo sa baga ay mahigpit na limitado at ang venous na dugo mula sa kaliwang aortic arch ay pinalalabas sa pamamagitan ng panizza foramen papunta sa kanang aortic arch, kung saan ito naghahalo.



Ang istraktura ng puso ng isang buwaya (RV - kanang ventricle, LV - kaliwang ventricle, FP - butas ng Panizza, RA - kanang aorta, LA - kaliwang aorta, PA - pulmonary aorta)
// pharyngula.org/Gazeta.Ru
kanin. Arterial system ng buwaya.
Sa mga reptilya, ang cellular na materyal ng sinus venosus (gumaganap sa mga amphibian) ay isinama sa atrial wall. Ang masa ng nodular tissue sa mga ibon at mammal ay bumubuo ng sinus node. Ito ay matatagpuan sa tagpuan ng superior vena cava sa kanang atrium. Ang knot tissue, na matatagpuan lamang sa puso, ay pinasisigla at kinokontrol ang mga contraction ng puso. Ito ay may ilang mga katangian ng parehong kalamnan at nervous tissue. Sa mga regular na pagitan, ang isang alon ng contraction ay kumakalat mula sa sinus node sa pamamagitan ng atrial na kalamnan. Kapag naabot nito ang atrioventricular (atrioventricular) node, ang salpok ay ipinapadala sa pamamagitan ng bundle ng nodular tissue patungo sa ventricles.
Mga ibon
Ang mga ibon ay humiwalay mula sa isang karaniwang ninuno na may mga buwaya, kaya ang sistema ng sirkulasyon ng mga ibon ay nagpapakita ng isang tiyak na pagkakatulad sa mga buwaya. Ang hitsura ng isang apat na silid na puso sa mga ibon at mammal ay ang pinakamahalaga ebolusyonaryong kaganapan, salamat sa kung saan ang mga hayop na ito ay maaaring maging mainit ang dugo. Dapat pansinin na ang mga ibon ay may mas advanced na mga baga at dobleng paghinga kaysa sa mga reptilya, na nagbibigay sa katawan ng oxygen sa isang sapat na lawak. Ang ganitong progresibong pagbabago ng sistema ng paghinga ng mga ibon ay nagbigay sa kanila ng mainit na dugo. Para sa mga ibon, hindi na kailangan ang insolation at muling pamamahagi ng dugo sa mga sisidlan sa lupa at sa ilalim ng tubig. Ang kaliwang aortic arch sa mga ibon ay ganap na nabawasan. Ang puso ng mga ibon ay apat na silid, dalawang arterial trunks ang umalis mula sa puso: ang kanang aortic arch ay nagsisimula mula sa kaliwang ventricle, at ang karaniwang trunk ng pulmonary arteries - mula sa kanang ventricle. Kaya, ang pag-unlad ng morphophysiological (ayon kay A.N. Severtsov) ay nakamit hindi sa pamamagitan ng komplikasyon, ngunit sa pamamagitan ng pagpapasimple at rasyonalisasyon ng mga sistema ng respiratory at circulatory. Ang portal system ng mga bato sa mga ibon ay wala, ang suplay ng dugo sa mga bato ay isinasagawa ng mga arterya ng bato na umaabot mula sa dorsal aorta.

mga mammal
Sa mga ninuno ng mga mammal, ang aorta ng tiyan ay hindi nahahati sa tatlo, tulad ng sa mga reptilya, ngunit sa dalawang arterial trunks: ang karaniwang trunk ng aortic arches at ang karaniwang trunk ng pulmonary arteries. Ang puso ng mga mammal ay may apat na silid, tulad ng sa mga ibon. Ang ventricle ng puso ay nahahati ng isang vertical septum sa kaliwa (kung saan nagsisimula ang karaniwang puno ng aortic arches) at ang kanan (mula sa kung saan nagmula ang pulmonary trunk). Sa harap, ang karaniwang trunk ng aortic arches ay nahahati sa kaliwang aortic arch (patuloy sa dorsal aorta) at ang innominate artery, na nagpapatuloy sa kanang subclavian artery. Ang mga carotid arteries ay nagsisimula nang simetriko: ang kaliwa - mula sa kaliwang aortic arch, ang kanan - mula sa innominate artery.
Sa pag-unlad ng embryonic ng mga mammal at tao, ang mga pagbabago sa puso at pangunahing mga daluyan ng dugo ng mga klase ng ninuno ay nire-recapital (paulit-ulit). Ang puso ay inilatag sa mga unang yugto ng pag-unlad sa anyo ng isang hindi nakikilalang aorta ng tiyan, na, dahil sa baluktot, ang hitsura ng mga partisyon at mga balbula sa lumen, ay nagiging sunud-sunod na dalawa-, tatlo- at apat na silid. Gayunpaman, ang mga recapitulations dito ay hindi kumpleto dahil sa katotohanan na ang interventricular septum ng mga mammal ay nabuo nang iba at mula sa ibang materyal kumpara sa mga reptilya. Samakatuwid, maaari nating ipagpalagay na ang apat na silid na puso ng mga mammal ay nabuo batay sa tatlong silid na puso, at ang interventricular septum ay isang neoplasma, at hindi ang resulta ng karagdagang pag-unlad ng reptile septum. Kaya, ang isang paglihis ay nagpapakita ng sarili sa phylogenesis ng vertebrate na puso: sa panahon ng morphogenesis ng organ na ito sa mga mammal, ang mga maagang yugto ng phylogenetic ay na-recapital, at pagkatapos ay ang pag-unlad nito ay nagpapatuloy sa ibang direksyon, katangian lamang para sa klase na ito.
Ang isang detalyadong pag-aaral ng pag-unlad ng puso sa butiki at mga embryo ng pagong at ang paghahambing nito sa magagamit na data sa mga amphibian, ibon, at mammal ay nagpakita na ang pangunahing papel sa pagbabago ng isang tatlong-silid na puso sa isang apat na silid ay nilalaro. sa pamamagitan ng mga pagbabago sa gawain ng Tbx5 regulatory gene, na gumagana sa una na nag-iisang ventricular rudiment. Kung ang Tbx5 ay ipinahayag (gumagana) nang pantay-pantay sa buong usbong, ang puso ay may tatlong silid, kung sa kaliwang bahagi lamang ito ay may apat na silid.
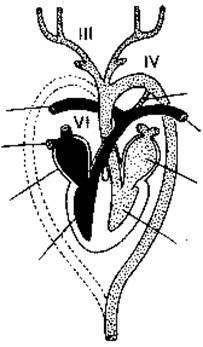

Lokasyon ng botallian duct (scheme): 1 - pulmonary artery; 2 - kanang sangay ng pulmonary artery; 3 - arko ng aorta; 4 - botalla (arterial) duct.
Ebolusyon ng mga organ system
"Wala sa biology
may katuturan kung hindi
tulad ng sa liwanag ng ebolusyon"
F.G. Dobzhansky
biyolohikal na ebolusyon- hindi maibabalik na proseso Makasaysayang pag-unlad buhay na kalikasan. Natutukoy sa pamamagitan ng pagkakaiba-iba, pagmamana at natural na pagpili. Sinamahan ng pagbagay ng mga organismo sa mga kondisyon ng pagkakaroon, ang pagbuo at pagkalipol ng mga species, ang pagbabago ng biogeocenoses at ang biosphere sa kabuuan.
Ebolusyon (mula sa lat. evolution - deployment) sa isang malawak na kahulugan - katulad ng pag-unlad; sa isang mas makitid, ang ideya ng mabagal, unti-unting pagbabago. Ang ebolusyon ay isang mahalagang sandali sa istruktura at mekanismo ng uniberso. Kaugnay nito, ang ebolusyon ng walang buhay na kalikasan, biyolohikal at sosyo-kultural na anyo ng ebolusyon ay nakikilala. Ang ebolusyonaryong pananaw sa mundo ay nagpapahintulot sa amin na makita ang pangkalahatan sa mga detalye at pagkakaisa sa pagkakaiba-iba.
Sa biology, ang pag-unlad ay nauunawaan bilang ang proseso ng malapit na magkakaugnay na quantitative (paglago) at qualitative (differentiation) na pagbabago ng mga indibidwal mula sa sandali ng pagsisimula hanggang sa katapusan ng kanilang buhay (ontogeny), gayundin sa buong buhay ng kanilang mga species sa Daigdig (historical development o phylogeny) .
Ang ebolusyon ay isang matatag na itinatag na siyentipikong katotohanan. Nagbibigay ito ng kahulugan, lohika at pagkakaisa sa buong napakalaking hanay ng kaalaman na naipon ng biology. Ang isang biologist ay maaaring lumihis mula sa ebolusyon sa loob lamang ng maikling sandali, na pinag-aaralan, halimbawa, ang istraktura ng dalawampu't unang pares ng mga paa sa centipedes, intercellular contact, ang istraktura ng isang nephron ng bato ng tao, isang neuron ng CNS, o iba pang mga istruktura. Gayunpaman, ang diskarte sa ebolusyonaryo (kasaysayan) ay nakakatulong upang malaman kung paano lumitaw ang kumplikadong istruktura at functional na mga yunit ng mga organo at ang mga organo mismo, pati na rin ang mga bagong uri ng mga organismo. Kung walang ebolusyonaryong pananaw sa mundo, ang biology ay nagiging isang koleksyon ng walang kahulugan, nakakalat, hindi maipaliwanag na siyentipikong data. Ang kaalaman sa ebolusyon ng hayop ay kinakailangan upang maunawaan ang mga sanhi ng maraming sakit at malformations. Ang impormasyong nakuha mula sa pag-aaral ng mga organismo sa iba't ibang antas ng pag-unlad (mga virus, bakterya, protista, worm, insekto, isda at iba pang hindi gaanong kumplikadong mga organismo) ay nagbibigay-daan sa amin upang mas maunawaan kung paano gumagana ang mga organo ng tao, kung ano ang nagiging sanhi ng mga sakit sa atin, at kung ano ang bago. mga pamamaraan na maaari nating gamitin upang mapabuti ang kalusugan at pahabain ang ating buhay.
Mode ng ebolusyon ng gene
Ang pinakamahalaga sa pagtuturo ng ebolusyon ay ang tanong kung paano umaangkop ang mga organismo sa panlabas na kapaligiran. Tinutulungan ng bioinformatics na masagot nang tama ang kardinal na tanong na ito. Sinusuri ng mga siyentipikong nagtatrabaho sa makabagong larangan ng genetics ang mga nucleotide sequence sa mga gene at ang pagkakasunud-sunod ng mga amino acid sa mga protina. Batay sa data ng bioinformatics, iminungkahi ng biologist ng Hapon na si Motto Kimura ang teorya ng molecular evolution. Ayon sa kanyang (malawakang tinatanggap) na mga pananaw, mayroong tatlong mga mode ng akumulasyon ng mga solong pagpapalit sa mga gene.
Ang una ay ang neutral na ebolusyon, kapag ang mga mutasyon ay hindi nagbabago sa istraktura o function ng gene at ang kaukulang protina. Ang mga ito ay katangian ng mga rehiyon ng DNA na hindi masyadong mahalaga para sa kaligtasan ng organismo at hindi nakakaapekto sa fitness nito. (Ang mga nasabing seksyon, halimbawa, ay mga hindi nagbibigay-kaalaman na mga seksyon ng isang gene - mga intron, mga intergenic na pagkakasunud-sunod - mga spacer). Sa kurso ng ebolusyon, ang mga naturang mutasyon ay naiipon sa mga genome sa humigit-kumulang na pare-pareho ang bilis, kaya nagsisilbi silang pamantayan ng oras sa ebolusyon ng molekular. Ang proseso ng akumulasyon ng naturang mga pagpapalit ay tinawag pa nga na isang molekular na orasan sa pamamagitan ng pagkakatulad sa isang atomic na orasan - ang pamantayan ng oras sa pisika. (Gamit, halimbawa, ang "molecular clock", tinukoy ng mga siyentipiko ang oras ng pagkakaiba-iba ng mga sanga mula sa parehong ninuno humahantong sa mga chimpanzee at tao).
Ang pangalawang mode ay ang pagpapatatag ng pagpili, sa ilalim ng impluwensya ng kung saan ay karaniwang partikular na mahalagang mga gene o ang kanilang mga seksyon; halos anumang pagpapalit sa kanila ay makakasira sa produkto ng gene at makakabawas sa fitness ng organismo. Ang rate ng akumulasyon ng mga mutasyon sa naturang pagpili ay mas mababa, mas mahalaga ang gene at mas mababa ang function nito ay direktang nauugnay sa panlabas na kapaligiran. Sa ilalim ng impluwensya ng pag-stabilize ng pagpili, kadalasang mayroong mga gene na "housekeeping" na nag-encode ng energy metabolism enzyme proteins, histone proteins, at marami pang iba. iba pa
Ang ikatlong paraan ng ebolusyon ay pagpili ng motibo. Ito ay nailalarawan sa pamamagitan ng isang pagtaas ng rate ng akumulasyon ng mga mutasyon, dahil sa ilalim ng mga bagong kondisyon ay pinapataas nila ang kakayahang umangkop ng organismo, na nagbibigay sa mga gene ng ilang mga bagong katangian (halimbawa, ang paglaban ng mga microorganism sa pagtaas ng antibiotic, mga insekto sa mga insecticides.
Sa ebolusyonaryong pagtuturo, 4 na pangunahing direksyon ng ebolusyonaryong pag-iisip ang nakikilala: 1) Lamarckism (ang posibilidad na magmana ng mga nakuhang katangian, ibig sabihin, pagkilala sa paglipat ng impormasyon mula sa phenotype patungo sa genotype. Ang pangalan ng doktrina ay nagmula sa pangalan ng Pranses siyentipiko na si Jean Baptiste Lamarck; mga impluwensya sa kapaligiran, lalo na sa mga unang yugto ng embryonic). Ang doktrina ay nagtataglay ng pangalan ng French naturalist na si Etienne Geoffroy Saint-Hilaire; 3) Darwinismo (ebolusyon sa pamamagitan ng natural na pagpili random na pagbabago). Ang teorya ay binuo ni Charles Darwin. 4) nomogenesis (pag-unlad batay sa mga regularidad, hindi aksidente). Ang may-akda ng konsepto ay si Lev Semenovich Berg. Ang mga tagapagtatag ng mga turong ito ay karaniwang tama, ngunit sa ilang mga paraan ang bawat isa sa kanila ay mali, marahil dahil sa pagkilala sa ganap na katotohanan.
Ang teorya ni Darwin ay nagbigay ng mga sagot sa mga pangunahing katanungan ng biology, kabilang ang kung bakit ang morpolohiya, pisyolohiya at pag-uugali ng mga organismo ay tumutugma sa kanilang mga kondisyon sa pamumuhay. Ang mga turo ni Darwin ay direktang nauugnay sa medisina. Pagkatapos ng lahat, ang gamot ay isang kumplikadong mga agham na nag-aaral ng isang tao na hindi lamang may sakit, ngunit malusog din - isang nilalang na karaniwang biological, umaasa sa kalikasan at sa parehong oras ay bahagi nito.
Ebolusyon ng sistema ng sirkulasyon ng mga vertebrates
Sa pagitan ng mga bahagi at organo ng katawan ng anumang organismo sa isang estado ng aktibong buhay, mayroong patuloy na pamamahagi ng mga sangkap at gas. Ang transportasyon ng isang likido at ang mga sangkap na nakapaloob dito ay maaaring mangyari batay sa osmosis, dahil sa pagsasabog, mga molekula ng carrier at mga espesyal na selula. Sa lubos na organisadong mga organismo, bilang karagdagan sa mga prosesong ito, ang isang epektibong paraan ng paglipat ng mga likido ay lumitaw, na isinasagawa ng mekanikal na gawain ng mga espesyal na "mga bomba sa puso". Ang mga Vertebrates ay may tatlong natatanging cavity na naglalaman ng mga likido: isang sistema ng mga daluyan ng dugo, isang pangalawang lukab ng katawan (ang kabuuan), at mga intercellular space na puno ng interstitial (tissue) fluid. (Ang terminong "likido" ay hindi ganap na tama, dahil ito solusyon sa koloid, ngunit sa pang-agham at pang-edukasyon na panitikan ito ay naayos nang mahabang panahon, at sa halip mahirap palitan ito ng isa pa, mas sapat na termino). Ang pag-agos ng likido mula sa mga intercellular space sa mga vertebrates ay nangyayari sa pamamagitan ng mga lymphatic vessel, na bulag na nagsisimula sa mga tisyu at dumadaloy sa anyo ng mga duct sa malalaking daluyan ng dugo.
ang pinakamahalagang mahalaga bahagi Ang circulatory system ay ang circulatory system. Ang sistema ng sirkulasyon ay gumaganap ng maraming mga pag-andar. Nagbibigay ito ng homeostasis at integridad ng katawan sa pamamagitan ng pagdadala ng oxygen. sustansya sa lahat ng mga organo, at carbon dioxide sa mga organ ng paghinga, naghahatid ng mga hormone sa mga target na selula, nakikilahok sa muling pamamahagi ng init sa katawan. Kasama ng transportasyon ng mga sangkap at gas, ang dugo ay gumaganap ng isang proteksiyon na function sa anyo ng mga immune reaction. Ang paglipat ng mga selula (kabilang ang mga selula ng kanser) ay nangyayari sa pamamagitan ng mga daluyan ng dugo at lymphatic.
Ang circulatory system ay naroroon sa iba't ibang uri ng non-chordates (nemerteans, annelids, mollusks, arthropods, echinoderms) at chordates. Chordates - superior uri, nahahati sa tatlong subtype: non-cranial, larval-chordal at cranial, o vertebrates. Ang circulatory system ng chordates ay nabuo sa isang espesyal na paraan at anuman ang estado ng circulatory system ng mga non-chordates. Ang pagpapabuti nito ay naganap sa pamamagitan ng pagkakaiba-iba (hitsura ng mga bagong istruktura), pagsasama-sama (hitsura ng mga bagong katangian ng system) at pagtindi ng mga pag-andar sa kurso ng phylogenesis (historikal na pag-unlad) ng mga chordates.
Sa aspetong medikal, ang ebolusyon ng sistema ng sirkulasyon sa mga non-cranial at vertebrate na hayop ay interesado, dahil pinapayagan tayong maunawaan ang sanhi ng atavistic (atavism - isang pagbabalik sa mga ninuno) malformations ng puso at mahusay na mga sisidlan. Dapat itong bigyang-diin na ang sistema ng sirkulasyon ay nabuo sa kasaysayan na may malapit na koneksyon sa iba pang mga sistema (nervous, respiratory, excretory, digestive) at ang pagsasaalang-alang nito, nang hiwalay sa iba pang mga sistema, ay dahil lamang sa mga pagsasaalang-alang sa didactic (upang mapadali ang pang-unawa. materyal na pang-edukasyon). Sa proseso ng ebolusyon, ang mga umiiral na organo ay naging mas kumplikado at ang mga bago ay nabuo (halimbawa, mga glandula ng endocrine), nagbago ang istraktura ng mga capillary, at lumitaw ang mga bagong regulatory at structural na mga molekula ng protina. Sa dugo ng tao, mayroong higit sa tatlong libong iba't ibang mga molekula ng protina.
Ito ay inilalagay sa embryo at nagpapatuloy sa mga matatanda sa mas malaki o mas maliit na lawak.
Ang gitnang sistema ng nerbiyos, kabilang ang utak, ay mahusay na binuo.
Ang iba't ibang grupo ng mga organismo ay nakararami sa pagbuo ng iba't ibang mga organo ng pandama, na umaabot sa kamag-anak na pagiging perpekto sa pag-unlad.
Ang panloob na balangkas ay mahusay na tinukoy. Ang nauunang dulo ng axial skeleton ay binago sa isang bungo.
Ang digestive tract ay nahahati sa oral cavity, pharynx, foregut (esophagus at tiyan), midgut (maliit na bituka), hindgut (malaking bituka).
Ang mga hasang slits ay kakaunti.
Ang mga kumplikadong organ sa paghinga ay bubuo.
at ang self-regulation ay ang mga bato.
Isang closed circulatory system, mayroong muscular organ para sa pagbomba ng dugo - ang puso.
Mga pulang selula ng dugo - ang mga pulang selula ng dugo ay naglalaman ng hemoglobin.
Ang mga organo ng hematopoiesis at blood depot ay ang pali.
Dalawang pares ng mga paa ang nabuo.
Ang lahat ng mga kinatawan ng subtype ay humantong sa isang aktibong pamumuhay. Kasama sa pangkat na ito ang: isda, amphibian reptile, ibon at mammal.
payat na isda
Ang mga integument ng katawan ay ctenoid o cycloid scales.
Lateral line - nakikita ang direksyon at lakas ng daloy ng tubig.
Ang organ ng pangitain - ang mga mata ay may isang spherical lens at isang flat cornea, nakikilala nila ang hugis at kulay, walang mga eyelid.
Ang organ ng pandinig - mayroon lamang panloob na tainga, ang gitna at panlabas ay wala.
Amoy - ang mga bulag na bag ay umaabot mula sa mga butas ng ilong, ang pang-amoy ay napaka banayad.
Pag-aanak - magkahiwalay na kasarian, ang babae ay nag-spawn sa tubig, at ang lalaki ay tinatakpan ito ng buto (gatas), i.e. Ang pagpapabunga ay panlabas, mayroong mga yugto ng larva.
Ang balangkas ay halos ganap na payat, may mga seksyon ng puno ng kahoy at buntot, ang gulugod ay nabuo ng bone vertebrae na may mga proseso, ang spinal canal. Bony ang bungo, na binubuo ng braincase, jaw bones, gill arches at gill covers.
Mga kalamnan - naka-segment at kinakatawan ng mga kalamnan ng puno ng kahoy na hugis-Z, na pinaghihiwalay ng connective tissue septa.
Sistema ng nerbiyos - binubuo ng utak at spinal cord at mga nerbiyos na umaabot mula sa kanila. Ang utak ay binubuo ng limang seksyon. Sa pag-uugali ng isda, ang mga unconditioned at conditioned reflexes ay ipinahayag.
Ang sistema ng sirkulasyon ay sarado at binubuo ng dalawang silid na puso at mga daluyan ng dugo. Isang bilog ng sirkulasyon ng dugo.
- oral cavity, pharynx, esophagus, tiyan, bituka na may bulag na proseso, atay, gallbladder, pancreas, swim bladder, anus.
Sistema ng paghinga - apat na kumpletong hasang at isang hanay ng mga filament ng hasang sa loob ng takip ng hasang.
Ang excretory system ay mahabang ribbon-like na mga bato, kung saan ang mga ureter ay umaabot, na pinagsama sa isang hindi magkapares na kanal ng urogenital papilla.
Mga amphibian
Integuments ng katawan - ang balat ay hubad, nakikilahok sa paghinga, ang mga glandula ng balat ay patuloy na naglalabas ng moisturizing mucus.
Lateral line - sa larvae at tailed amphibians.
Ang organ ng paningin - ang mga mata ay may mga talukap ng mata, isang matambok na kornea, isang lenticular lens.
Ang organ ng pandinig ay ang panloob, gitnang tainga na may tympanic membrane, sa loob ng auditory ossicle, stirrup.
Pang-amoy - panlabas na butas ng ilong na humahantong sa mga kapsula ng olpaktoryo.
Ang lasa ay mga sensitibong selula (sa bibig at sa buong katawan).
Pagpaparami - dioecious, ang mga babae ay naglalagay ng kanilang mga itlog sa tubig, at ang mga lalaki ay tinatakpan ito ng isang buto, i.e. Ang pagpapabunga ay panlabas, ang larvae ay kahawig ng pritong isda.
Skeleton - ang gulugod, bungo at balangkas ng mga limbs at ang kanilang mga sinturon. Wala ang dibdib.
Ang mga kalamnan ay makapangyarihan at kumplikadong mga kalamnan ng mga paa.
Sistema ng nerbiyos - 10 pares ng nerbiyos ang umaalis dito.
Ang sistema ng sirkulasyon ay sarado, ang puso ay tatlong silid, binubuo ng dalawang atria at isang ventricle. Dalawang bilog ng sirkulasyon ng dugo.
Digestive system - oral opening, oropharyngeal cavity, esophagus, sa oral cavity, dila, tiyan, bituka, pancreas, malaking atay, gallbladder, bituka ay nahahati sa manipis at makapal, undigested residues - sa pamamagitan ng cloaca.
Sistema ng paghinga - huminga sa pamamagitan ng mga baga at balat, paghinga ng hasang sa yugto ng embryonic.
Sistema ng excretory - kidney excretory ducts ureters bumubukas sa cloaca.
Mga ibon
Integuments ng katawan - ang katawan ay natatakpan ng mga balahibo, malibog na mga kalasag sa mga binti.
Lateral na linya - hindi.
Ang organ ng pangitain - ang mga mata ay nilagyan ng mga talukap ng mata, kasama. at ang ikatlong takipmata - ang nictitating lamad, ay may pambihirang pagbabantay, nakikilala ng mabuti ang mga kulay.
Ang organ ng pandinig ay ang panloob, gitna at panlabas na tainga, nakakarinig sila ng maayos.
Ang pang-amoy ay ang lukab ng ilong, ang pang-amoy ay hindi gaanong nabuo.
Panlasa - mahinang nabuo (mga sensitibong selula sa bibig).
Pagpaparami - dioecious. Ang pagpapabunga ay panloob. Oviparous. Ang mga itlog ay naglalaman ng isang pula ng itlog at natatakpan ng isang calcareous shell. Walang mga yugto ng larva.
Skeleton - ang mga manipis na buto na may malawak na mga cavity ng hangin ay nagbibigay ng liwanag sa balangkas, lakas dahil sa pagsasanib ng isang bilang ng mga buto, ang gulugod, bungo, balangkas ng mga limbs at sinturon ng paa.
Mga kalamnan - ang mga kalamnan ng pectoral ay makapangyarihan, itinaas at ibababa ang mga pakpak, ang mga kumplikadong kalamnan ng mga hind limbs.
Ang sistema ng nerbiyos ay mas kumplikado kaysa sa amphibian at reptilya. Ang masa ay katumbas ng masa ng spinal, forebrain hemisphere, na nauugnay sa pagpapalawak ng aktibidad ng motor at ang komplikasyon ng pag-uugali. Ang cerebellum ay napakalaki, ito ay dahil sa kumplikadong paggalaw ng mga ibon.
Ang sistema ng sirkulasyon ay isang kumpletong paghihiwalay ng malaki at maliliit na bilog ng sirkulasyon ng dugo. Ang arterial at dugo ay hindi naghahalo, ang puso ay may apat na silid, malaki ang dami, ay binubuo ng dalawang atria at dalawang ventricles, ang puso ay gumagana nang masinsinan.
Digestive system - oral cavity, dila, salivary glands, mahabang esophagus na may goiter, thin-walled two-chamber na tiyan, glandular section, toasty-walled muscular section. Manipis at makapal ang bituka. Ang manipis ay naglalaman ng pancreas, ang makapal ay pinaikli at nagtatapos sa isang cloaca.
Respiratory system - pulmonary breathing, ang trachea ay nahahati sa dalawa, ang mas mababang isa ay may vocal cords. Ang mga baga ay may may ngipin na istraktura, mga air sac.
Sistema ng excretory - bato, ureter, cloaca. Walang pantog.
mga mammal
Integuments ng katawan - ang katawan ay natatakpan ng lana - buhok, bukod sa kung saan mayroong mga tactile hair - vibrissae; dalawang uri ng mga glandula ng balat: sebaceous at pawis.
Lateral na linya - hindi.
Ang organ ng pangitain - ang mga mata ay nilagyan ng mga eyelid, ngunit ang ikatlong takipmata ay pasimula, ang visual acuity ay nakasalalay sa mga kondisyon ng pamumuhay. Ang paningin ng kulay ay mas mahina kaysa sa mga ibon, o wala.
Ang organ ng pandinig - mayroon silang panloob, gitna at panlabas na tainga, kadalasan ay mahusay na binuo ang pandinig, ang ilan ay may tunog na lokasyon.
Ang pang-amoy ay ang lukab ng ilong na may mga concha ng ilong, na natatakpan ng epithelium, ang pang-amoy ay banayad.
Panlasa - mga bato, sensitibong mga selula, na nahuhulog sa ibabaw ng dila sa anyo ng mga papillae.
Pagpaparami - dioecious. Ang pagpapabunga ay panloob. Ang embryo, bilang panuntunan, ay bubuo sa katawan ng isang babae, viviparous.
Ang balangkas ay isang perpektong balangkas, mga subdibisyon: katawan at ulo (bungo), balangkas ng mga paa at sinturon ng paa. Ang vertebrae ay may mga patag na ibabaw kung saan matatagpuan ang mga cartilaginous disc. Malaking brain box.
Mga kalamnan - naiiba at kinakatawan ng maraming magkakaibang lokasyon na mga kalamnan, ang pagkakaroon ng diaphragm ay katangian, ang mga subcutaneous na kalamnan ay binuo, gayahin sa mukha.
Ang nervous system - ang utak ay nailalarawan sa pamamagitan ng malaking sukat. Lumalaki ang cerebral cortex. Sa karamihan ng mga species, hindi ito makinis, ngunit bumubuo ng mga furrow na nagpapataas ng lugar nito. Ang malaking cerebellum ay naiba sa mga seksyon.
Ang sistema ng sirkulasyon ay kumplikado. Dalawang bilog ng sirkulasyon ng dugo, perpektong mga sisidlan, aorta, mga capillary.
Ang sistema ng pagtunaw ay kumplikado digestive tract mahaba. Pre-oral cavity (ngipin, pisngi, panga), oral cavity, ngipin, dila, pharynx, esophagus, tiyan na may mga glandula, bituka, makapal at manipis. Glandular system: atay, gallbladder, pancreas. Nagtatapos sa anus.
Sistema ng paghinga - mga baga, trachea at bronchi, na mahusay na binuo. Sa rehiyon ng mga baga, ang bronchi ay nahahati sa isang malaking bilang ng mga sanga. Ang pinakamaliit ay nagtatapos sa alveoli - mga bula kung saan nangyayari ang palitan ng gas.
Sistema ng excretory - ang mga bato, na binubuo ng panlabas (cortical) at panloob (cerebral) na mga layer, ang mga ureters, ang pantog, ang urethra. Ang mga glandula ng pawis ay gumaganap ng bahagi ng excretory function.
1.1. Pinagmulan at pag-andar ng integument ng katawan.
Ang epidermis ng integument ng katawan ay nabuo mula sa ectoderm, at ang connective tissue ng balat ay nabuo mula sa mesoderm.
Ang mga integument ng katawan ay gumaganap ng mga sumusunod na function:
Proteksyon mula sa mga nakakapinsalang impluwensya sa kapaligiran;
Pagdama ng stimuli;
Pakikilahok sa metabolismo;
Ang integument ng katawan ay isang karagdagang organ ng paghinga at paglabas, at sa mga arthropod ito ang panlabas na balangkas.
1.2. Ebolusyon ng mga pantakip sa katawan sa mga invertebrates.
Ang mga integument ng katawan sa mga invertebrates ay pangunahing nabuo ng ectoderm at mga derivatives nito. Kaya, ang skin-muscular sac ng flatworms ay binubuo ng tisyu ng integumentaryo- tegument, na isang multinuclear non-cellular na istraktura at tatlong layer ng mga kalamnan - longitudinal, transverse at dorsoventral. Ang mga galaw na ginagawa nila ay mabagal at hindi perpekto. Sa mga turbellarian, ang katawan ay natatakpan ng cilia na kasangkot sa paggalaw.
Balat-muscular sac mga bulate ay binubuo ng isang multi-layered, elastic at bahagyang extensible cuticle, hypodermis, na isang solong cytoplasmic mass, hindi nahahati sa mga indibidwal na mga cell at naglalaman ng isang malaking bilang ng mga nuclei, at isang layer ng longitudinal at makinis na mga kalamnan. Ang cuticle ay pangunahing gumaganap ng isang proteksiyon na function. Ang mga kalamnan ay matatagpuan sa anyo ng dalawang longitudinal strands - sa ventral at dorsal side ng katawan. Ang kanilang kahaliling pag-urong ay nagbibigay ng masiglang pagbaluktot at mga paggalaw ng extension at mabilis na paggalaw ng katawan sa espasyo.
Ang katawan ng mga annelids ay natatakpan ng isang skin-muscle sac, na binubuo ng isang layer ng epithelium at dalawang layers ng mga kalamnan: ang panlabas na layer ng annular at ang panloob na layer na nabuo sa pamamagitan ng longitudinal muscle fibers.
Kaya, ang ebolusyon ng mga integument ng katawan sa mga invertebrates ay nagpatuloy mula sa ciliated epithelium ng turbellarians, i.e. ciliary worm, na gumaganap ng function ng primitive organo ng paggalaw sa kanila, sa flat, wala ng ciliated outgrowths epithelium ng flukes, tapeworms, bilog at annelids.
Ang mga integument ng katawan ng mga arthropod ay kinakatawan ng isang chitinized cuticle na nagpoprotekta sa mga organo mula sa mga panlabas na nakakapinsalang impluwensya at isang panlabas na balangkas kung saan nakakabit ang mga nakahiwalay na kalamnan, na binubuo ng mga striated na fibers ng kalamnan.
1.3. Ebolusyon ng mga pantakip sa katawan sa mga chordates.
Sa chordates, mayroong dalawang layer sa balat: ang itaas ay ang ectodermal epidermis at ang mas mababang isa ay ang mesodermal corium o ang balat mismo, o ang dermis.
Ang ebolusyon ng balat sa mga chordates ay sumunod sa landas ng paglipat mula sa isang solong-layer na epithelium patungo sa isang multi-layer, pati na rin ang isang pagbabago sa ratio sa pagitan ng mga layer patungo sa isang mas malaking pag-unlad ng balat mismo.
Para sa mga mas mababang chordates, lalo na para sa mga hindi cranial, isang mahinang antas ng pagkita ng kaibhan ng parehong mga layer ng balat ay katangian. Ang epidermis ay single-layer cylindrical, na nakatali sa labas ng manipis na porous cuticle na naglalaman ng unicellular mucous glands. Sa ilalim ng epidermis ay namamalagi ang isang manipis na base plate, at sa ilalim nito ay isang mahinang layer ng gelatinous connective tissue, na kumakatawan sa aktwal na balat o dermis.
Sa mas mataas na chordates, i.e. sa vertebrates, ang integuments ng katawan ay hindi binubuo ng isang solong layer, ngunit ng isang multi-layered ectodermal epidermis at isang mesodermal fibrous corium, o skin proper. Ang epidermis ay may dalawang layer. Ang mga cell ng mas mababang, tinatawag na cambial, layer ay nagsisilbi upang magparami ng mga bagong layer ng mga cell, at ang mga cell ng itaas na layer ay may proteksiyon na function. Ang itaas na layer ay may maraming mucous at iba pang mga glandula; sa karamihan ng mga terrestrial na hayop, malibog na mga scute, kaliskis, balahibo, buhok, kuko, claws, sungay, hooves, atbp ay nabuo dito dahil sa keratinizing cell. Sa itaas na layer ng epidermis, ang mga cell ay nag-iiba, namamatay, at nag-desquamate.
Kabanata 2
Ang ebolusyon ng balangkas.
2.1. Pinagmulan at pag-andar ng balangkas ng hayop.
Ang mga pormasyon ng suporta sa mga invertebrate, na kumakatawan sa kanila ng isang permanenteng hugis ng katawan, ay napaka-magkakaibang. Ang mga ito ay ecto-, ento- at mesodermal na pinagmulan. Sa mga vertebrates, ang balangkas ay pangunahing mesodermal ang pinagmulan.
Ang balangkas sa katawan ng hayop ay gumaganap ng iba't ibang mga pag-andar:
Tinitiyak ang isang pare-parehong hugis ng katawan;
Passive na bahagi ng musculoskeletal system;
Proteksyon laban sa mekanikal at iba pang mga impluwensya;
hematopoietic function.
2.2. Ang ebolusyon ng balangkas sa isang serye ng mga invertebrates.
Sa mga espongha, ang mga sumusuportang pormasyon ay kinakatawan ng mga karayom na may ibang komposisyon ng kemikal.
Sa mga lukab ng bituka, lumilitaw ang isang siksik na sumusuporta sa plato (mesogley), na sumasakop sa isang lugar sa pagitan ng ecto- at endoderm. Ang balangkas ng mga coral polyp ay bubuo mula sa ectoderm. Sa mga arthropod, ang panlabas na balangkas ay kinakatawan ng isang chitinous na takip, na kinabibilangan ng mga pag-andar ng proteksyon laban sa mekanikal na pinsala at ang panlabas na balangkas, kung saan ang mga striated na kalamnan na unang lumitaw sa mga arthropod ay nakakabit.
Bivalves at mga gastropod may mga shell na nabuo sa pamamagitan ng mga pagtatago ng mantle. mga cephalopod may mga kumplikadong cartilaginous formations na nagpoprotekta sa mga nerve center at sensory organ.
2.3. Ebolusyon ng kalansay sa mga chordates.
Tulad ng mga invertebrates, ang skeleton ng chordates ay gumaganap ng tungkulin ng pagprotekta sa mga organo at nagsisilbing suporta para sa mga organo ng paggalaw.
Axial skeleton dumaan sa malalaking pagbabago sa kurso ng ebolusyon.
Sa mas mababang chordates, ang notochord ay ang axial skeleton, habang sa mas mataas na mga ito ay unti-unting pinapalitan ng pagbuo ng vertebrae. Sa vertebrae, ang katawan, itaas at mas mababang mga arko ay nakikilala.
Kaya sa mga cyclostomes, ang notochord ay nagpapatuloy sa buong buhay, ngunit lumilitaw ang mga vertebral anlages, na mga maliliit na cartilaginous formations na metamerically na matatagpuan sa itaas ng notochord. Ang mga ito ay tinatawag na upper arches.
Sa primitive na isda, bilang karagdagan sa itaas na mga arko, lumilitaw ang mas mababang mga arko, at sa mas mataas na isda, lumilitaw din ang mga vertebral na katawan. Ang mga vertebral na katawan sa karamihan ng mga isda at mas matataas na hayop ay nabuo mula sa tissue na nakapalibot sa notochord, pati na rin mula sa mga base ng mga arko. Ang itaas at ibabang mga arko ay nagsasama sa mga vertebral na katawan. Ang mga dulo ng itaas na mga arko ay lumalaki nang magkasama upang bumuo ng spinal canal. Sa mas mababang mga arko, lumilitaw ang mga proseso, kung saan nakakabit ang mga tadyang. Ang isda ay may dalawang seksyon ng gulugod - puno ng kahoy at buntot. Ang mga labi ng notochord sa isda ay napanatili sa pagitan ng mga vertebral na katawan.
Sa mga amphibian, sa mga unang yugto ng pag-unlad, ang notochord ay pinalitan ng gulugod. Mayroon nang apat na seksyon sa gulugod: cervical, thoracic, sacral at caudal. Ang cervical region ay mayroon lamang isang vertebrae, habang ang thoracic region ay may limang vertebrae. Ang mga maliliit na tadyang ay nakakabit sa thoracic vertebrae, na malayang nagtatapos. Ang sacral region, tulad ng cervical, ay may kasamang isang vertebra, na siyang suporta para sa mga buto ng pelvis at hind limbs. Ang seksyon ng buntot sa mga amphibian na walang buntot ay nagsasama sa isang buto, at sa mga caudates ay binubuo ito ng isang malaking bilang gulugod.
Ang mga reptilya ay may limang seksyon sa gulugod: cervical, thoracic, lumbar, sacral, at caudal. Sa rehiyon ng servikal, ang iba't ibang uri ng mga reptilya ay may ibang bilang ng vertebrae, ngunit ang pinakamalaki ay walo. Ang unang vertebra ay tinatawag na atlas at may hugis ng singsing, at ang pangalawa ay tinatawag na epistrophy at may prosesong odontoid kung saan umiikot ang unang vertebra. Sa rehiyon ng thoracic, ang bilang ng mga vertebrae ay hindi pare-pareho; ang mga buto-buto ay nakakabit sa kanila, karamihan sa mga ito ay konektado sa sternum, na bumubuo ng dibdib sa unang pagkakataon sa mas mataas na mga hayop. Mayroon lamang 22 vertebrae sa thoracolumbar region, at dalawa sa sacral region. Ang mga buto-buto ay nakakabit din sa vertebrae ng lumbar at sacral na rehiyon. Sa rehiyon ng caudal, ang bilang ng mga vertebrae ay nag-iiba, kung minsan ilang dosena.
Sa mga ibon, ang vertebral column ay katulad ng sa mga reptilya, ngunit may isang tiyak na espesyalisasyon na may kaugnayan sa caudal. at ang gulugod - ang trunk ribs. mula sa YVDr na nakapalibot sa chord, pati na rin mula sa base ng pagbuo ng arko, na may paraan ng pamumuhay. Kasama sa cervical region ang hanggang 25 vertebrae, na nagbibigay ng higit na kadaliang kumilos.
Ang mga mammal ay may limang seksyon sa gulugod: cervical, thoracic, lumbar, sacral, at caudal. Mayroong pitong vertebrae sa cervical region, at isang variable na bilang ng vertebrae sa thoracic region (mula 9 hanggang 24, ngunit mas madalas 12-13). Ang mga buto-buto ay sumasali sa thoracic vertebrae, ang malaking bilang nito ay konektado sa sternum. Kasama sa rehiyon ng lumbar ang tatlo hanggang siyam na vertebrae. Ang sacral vertebrae ay pinagsama upang mabuo ang sacrum, at kasama ang caudal spine magkaibang halaga vertebrae sa iba't ibang mammalian species.
Balangkas ng ulo. Ang balangkas ng ulo ay ang bungo. Ito ay matatagpuan sa nauunang dulo ng balangkas at binubuo ng dalawang bahagi: ang cranium at ang visceral skeleton, na naiiba sa bawat isa kapwa sa pinagmulan at sa kanilang pag-andar. Ang cranium ay nagsisilbing lalagyan para sa utak, at ang visceral skeleton ay nagbibigay ng suporta para sa mga organo ng anterior na bahagi ng alimentary canal.
Sa proseso ng ebolusyon, ang pinakamalaking pagbabago ay nangyayari sa visceral na rehiyon. Sa mga embryo ng lahat ng vertebrates, at sa mga lower vertebrates sa buong buhay, ang visceral skeleton ay binubuo ng mga arko na sumasaklaw sa anterior na bahagi ng digestive tube. Sa isda, naiba sila sa arko ng panga (pagkuha ng pagkain), arko ng hyoid (kabit sa cranium), at arko ng hasang (kabit ng mga hasang).
Sa mga hayop sa terrestrial, ang visceral skeleton ay lubos na nagbabago at nabawasan: ang itaas na bahagi ng arko ng panga ay sumasama sa ilalim ng bungo, ang mga maliliit na buto ay nabuo mula sa hyoid arch, na bahagi ng gitnang tainga. Ang pangalawa at pangatlong branchial arch sa mga mammal ay bumubuo sa thyroid cartilage, habang ang pang-apat at ikalimang arko ay bumubuo sa natitirang mga cartilage ng larynx.
balangkas ng paa. Mayroong dalawang uri ng libreng paa. Ito ang mga palikpik ng isda at ang limang daliri na paa ng mga mammal. Ang limang-daliri na mga limbs ng vertebrates ay may isang napaka-magkakaibang istraktura, na nauugnay sa pagganap ng iba't ibang mga pag-andar ng mga ito. Halimbawa, ang mga burrowing limbs ng nunal, floating limbs ng seal, climbing limbs ng monkeys, etc. Ngunit gayunpaman, sa kabila ng pagkakaiba, ang mga limbs ng vertebrates ay nagpapanatili ng isang karaniwang plano sa istruktura, na nagpapatunay sa pagkakapareho ng kanilang pinagmulan.
Sa unang pagkakataon, lumitaw ang mga limbs sa isda at kinakatawan ng mga palikpik. Ang mga ito ay magkapares na pectoral at ventral fins. Sa karamihan ng mga isda, ang mga palikpik ay binubuo ng radial thin bony rays at nagsisilbing baguhin ang direksyon ng paglangoy, sa halip na suportahan ang katawan. Sa lobe-finned fish, ang pagpapalaki ng mga sinag ay binalak dahil sa pagsasanib at paggamit ng mga palikpik bilang suporta at paggalaw kasama ang solidong base ng mga natutuyong reservoir. Samakatuwid, ang mga palikpik ng mga sinaunang isda na may palikpik na lobe ay ang batayan para sa pag-unlad ng mga limbs ng mga vertebrates. Isang mahalagang katangian ang pagbabagong-anyo ng mga palikpik sa mga limbs ng terrestrial vertebrates ay ang kapalit ng isang malakas na koneksyon ng mga elemento ng kalansay na may isang movable na koneksyon sa anyo ng mga joints. Bilang isang resulta, ang paa ay naging isang kumplikadong movable lever, kung saan ang tatlong buto ay nakikilala: ang balikat, bisig at kamay. Mayroong dalawang sinturon sa paa - balikat at pelvic.
Dagdag pa, ang ebolusyon ng forelimb ay sumunod sa landas ng pagpapahaba ng balikat at bisig, pinaikli ang pulso, binabawasan ang bilang ng mga buto sa rehiyon ng carpal (sa mga amphibian - 3 mga hanay, sa mga mammal - 2 mga hilera) at pagpapahaba ng mga distal na seksyon, i.e. phalanges ng mga daliri.
Ang balangkas ng kamay ng tao ay nailalarawan din ng isang pangkalahatang plano sa istruktura na may mga forelimbs ng mga vertebrates, ngunit kasama nito, mayroon din itong mahahalagang pagkakaiba, dahil ang kamay ng tao ay hindi lamang isang sandata ng paggawa, kundi pati na rin ang resulta nito at may kakayahang ng pagsasagawa ng iba't ibang kilos.
2.4. Anomalya at malformations ng skeleton ng tao.
1. Ang pagkakaroon ng ribs sa lower cervical o sa unang lumbar vertebrae. Alinsunod sa ebolusyon ng mga vertebrates sa mga tao, sa panahon ng pag-unlad ng embryonic, ang mga buto-buto ay inilalagay sa lahat ng bahagi ng gulugod, ngunit pagkatapos ay napanatili lamang sila sa rehiyon ng thoracic, at sa ibang mga rehiyon ang mga buto-buto ay nabawasan. Ngunit kung minsan ang isang tao ay may katulad na mga atavism.
2. Pagkakaroon ng tail vertebrae. Sa panahon ng embryogenesis sa mga tao, tulad ng sa mga vertebrates, 8-11 tail vertebrae ay inilatag, pagkatapos ay sila ay nabawasan at 4-5 underdeveloped vertebrae ay nananatili, na bumubuo ng coccyx. Minsan may mga atavistic na palatandaan sa anyo ng pagkakaroon ng caudal spine.
3. spina bifida- Ito ay isang pangkaraniwang anomalya na nangyayari kapag may paglabag sa pagsasanib ng itaas na mga arko ng vertebrae. Madalas itong nagpapakita ng sarili sa lumbosacral na rehiyon ng gulugod at, depende sa lalim at lawak ng split, ay maaaring magkaroon ng iba't ibang antas ng kalubhaan.
4. Ang presensya sa tympanic cavity ng isang auditory bone lamang - column. Ang kaguluhan na ito, na tumutugma sa istraktura ng sound-transmitting apparatus ng mga amphibian at reptile, ay ang resulta ng hindi tamang pagkita ng kaibahan ng mga elemento ng jaw gill arch sa auditory ossicles. Ito ay isang recapitulation ng mga pangunahing yugto ng phylogenesis ng visceral skull sa ontogeny.
5. Heterotopia ng sinturon ng itaas na mga paa't kamay. Ito ang paggalaw ng sinturon ng itaas na mga limbs mula sa cervical region hanggang sa antas ng 1-2 thoracic vertebrae. Ang anomalyang ito ay tinatawag na Sprengel's disease o congenital high standing ng scapula. Ito ay ipinahayag sa katotohanan na ang sinturon ng balikat sa isa o magkabilang panig ay ilang sentimetro na mas mataas kaysa sa normal na posisyon. Ang mekanismo ng naturang paglabag ay nauugnay sa parehong paglabag sa paggalaw ng mga organo at isang paglabag sa morphogenetic correlations.
6. Polydactyly- ang resulta ng pagbuo ng mga anlages ng karagdagang mga daliri, katangian ng malayong mga anyong ninuno.
7. Flat feet, clubfoot, makitid rib cage, kawalan ng protrusion sa baba- atavistic anomalya ng skeleton, na madalas na matatagpuan at mga anobolism (extension) na lumitaw sa panahon ng phylogenesis ng primates.
Ang Vetulicolia, unang natagpuan sa Greenland noong 1911, ay matatagpuan sa mga deposito ng Cambrian ng iba't ibang rehiyon ng mundo - mula sa China hanggang Canada. Bulag, ngunit may malawak na bibig, maaari silang lumangoy salamat sa mga paggalaw ng buntot. Mula sa isang ekolohikal na pananaw, ang vetulicolia ay kahawig ng mga maliliit na whale shark, sinasala ang plankton at organikong bagay mula sa column ng tubig.
Sa ngayon, 14 na species ng mga hayop na ito ang kilala sa agham, ngunit dahil sa napakahirap na pangangalaga ng kanilang mga relasyon sa pamilya, pati na rin ang mga detalye ng morpolohiya at hitsura sa ngayon ay nanatiling hindi gaanong naiintindihan. Ito ay kilala lamang na ang katawan ng Vetulicolia ay malinaw na nahahati sa dalawang seksyon - isang guwang na anterior at isang naka-segment na posterior. Ang istraktura ng nauuna na seksyon ay nagmumungkahi na ang pinakamalapit na kamag-anak ng vetulicolia ay maaaring mga primitive chordates tulad ng mga salp at ascidian, at ang buntot ay nagbubunga ng mga saloobin ng mga arthropod. Nakatulong ang isang bagong pagtuklas upang maunawaan ang mga ugnayan ng pamilya ng grupo.
Kamakailan, natuklasan ng isang grupo ng mga paleontologist ng Australia na pinamumunuan ni Diego Garcia-Bellido ng University of Adelaide ang mga fossil ng bagong species ng vetulicolia sa Kangaroo Island. Pinangalanan ito ng mga siyentipiko na Nesonektris aldridgei, bilang parangal sa sikat na mananaliksik ng grupong ito, si Dick Eldridge mula sa British University of Leicester. Ang generic na pangalan ng hayop sa Greek ay nangangahulugang "manlalangoy sa isla".
Ang mga nesonectrises ay lumaki sa halos 13 cm ang haba. Ang isang kakaibang tampok ng kanilang pangangalaga ay ang mga buntot at mga seksyon sa harap ay madalas na matatagpuan nang hiwalay, iyon ay, hindi sila mahigpit na konektado at nadurog sa ilang sandali pagkatapos ng pagkamatay ng mga hayop. Ang mga paleontologist ay mas interesado sa tubo na tumatakbo sa kahabaan ng katawan tulad ng isang bituka: hinati ito ng mga partisyon sa magkakahiwalay na mga bloke.
"Ito ay ganap na hindi naaayon sa bituka (na isang guwang na tubo), ngunit angkop na angkop sa isang cartilaginous notochord (o notochord), sabi ni Garcia-Bellido. "Kaya, ngayon ay maaari tayong gumawa ng mga konklusyon tungkol sa eksaktong kung saan matatagpuan ang grupong ito sa puno ng buhay."
Ang notochord ay matatagpuan sa halos lahat ng vertebrates sa embryonic stages of development, kadalasang nagbibigay daan sa spine habang sila ay tumatanda. Ang ilang mga primitive chordates ay nagpapanatili ng notochord sa buong buhay nila, habang ang iba, tulad ng sea squirts, ay mayroon lamang nito sa mga unang yugto ng pag-unlad. Kung ang mga vetulicolian ay may notochord, kung gayon ay inilalagay sila sa grupo ng mga ninuno o kahit man lang "mga pinsan" ng lahat ng iba pang chordates, kabilang ang mga tao.
"Malapit silang kamag-anak ng mga vertebrates. Ang Vetulicolia ay may mahabang buntot na sinusuportahan ng isang matibay na baras na kahawig ng isang notochord, na siyang pasimula ng gulugod at isang natatanging katangian ng mga vertebrates at kanilang mga kamag-anak," sabi ng paleontologist ng Australia. Balak ngayon ni Garcia-Bellido at ng kanyang koponan na muling suriin ang mga kilalang labi ng iba pang mga vetulicolian upang subukang makahanap din ng mga notochords sa kanila. Bilang karagdagan, ang mga paghuhukay ay magpapatuloy sa Kangaroo Island, kung saan, salamat sa isang pambihirang hanay ng mga pangyayari, ang mga imprint ng malambot na mga tisyu ng mga hayop sa panahon ng Cambrian ay napanatili sa isang fossil na estado. 
Artikulo Isang bagong vetulicolian mula sa Australia at ang kaugnayan nito sa chordate affinities ng isang enigmatic Cambrian group na inilathala sa BMC Evolutionary Biology
Ang mga panga ng mga vertebrates ay pinaniniwalaang nag-evolve mula sa nauunang pares ng mga arko ng hasang ng kanilang pinakaunang mga ninuno na parang isda. Sa wakas, natagpuan ng mga siyentipikong Amerikano ang dokumentaryong ebidensya ng hypothesis na ito sa fossil ng organismong Cambrian na Metaspriggina.

Metaspriggina walcotti. Muling pagtatayo: Marianne Collins
Ayon sa modernong siyentipikong konsepto, ang mga panga at ilang maliliit na buto ng panloob na tainga ng tao ay sumusubaybay sa kanilang mga ninuno sa mga arko ng hasang - mga kakaibang istruktura ng kalansay na sumusuporta sa mga hasang ng primitive na parang isda na vertebrates noong Cambrian o kahit na Precambrian times. Gayunpaman, dahil sa hindi magandang pag-iingat at hindi kumpletong rekord ng geological, hindi posible hanggang kamakailan lamang na mahanap ang mga arko na ito sa kanilang lokasyon sa katawan ng isang organismo.
Inilarawan ni Propesor Simon Morris ng Unibersidad ng Cambridge sa UK at ng kanyang kasamahan na si Dr. Jean-Bernard Caron ng Unibersidad ng Toronto at ng Royal Ontario Museum ang mga istruktura ng hasang ng maagang chordate na Metaspriggina walcotti, na nabuhay 505 milyong taon na ang nakalilipas. Ang mga fossil ng kakaibang napreserbang metaspriggin na ito ay natagpuan noong 2012 sa sikat na lugar ng unang bahagi ng Cambrian fauna ng Burges Shale, kasama ang apat na dosenang iba pang fossil.
"Ang detalye sa mga fossil na ito ay napakaganda," sabi ni Morris. "Maging ang mga mata ay ganap na napanatili at perpektong nakikita." Ang dalawang malalaking mata ng metaspriggin ay nakalagay sa pinakadulo ng nguso kaya't tila nakaumbok. At hindi kalayuan sa kanila, nakita ng mga siyentipiko ang bahagyang hubog na mga baras - ang parehong mga arko ng gill na nakatakdang magbunga ng mga panga at pandinig na buto ng mga vertebrates.
"Ang mga arko na ito ay matagal nang kilala na gumaganap ng isang mahalagang papel sa ebolusyon ng mga vertebrates, kabilang ang paglitaw ng kanilang mga panga at maliliit na buto na kasangkot sa paghahatid ng tunog sa mga mammal," sabi ng propesor sa Britanya. "Hanggang ngayon, gayunpaman, ang kakulangan ng mga de-kalidad na fossil ay nangangahulugan na ang aming pag-unawa sa lokasyon ng mga arko na ito sa mga unang vertebrates ay puro hypothetical."
Ito ay lumabas na ang mga gill arches ay matatagpuan sa katawan ng hayop nang pares, na kung saan ay ang pinakamahusay na posibleng kumpirmasyon ng hypothesis ng pinagmulan ng isang pares ng mga panga mula sa kanila. Kasabay nito, ang unang pares ng mga arko sa Metaspriggina ay mas makapal din kaysa sa lahat ng iba pa, na, marahil, ang unang ebolusyonaryong hakbang sa daan patungo sa paglitaw ng mga lamok.
Pagkatapos ng karagdagang pagsusuri sa istraktura at lokasyon ng mga kalamnan ng metaspriggin, ang mga paleontologist ay dumating sa konklusyon na ang hayop na ito ay isang aktibo at mobile na manlalangoy, hindi mas mababa sa modernong trout. At ang isang pares ng malalaking mata at isang organ ng olpaktoryo ay nagpapahintulot sa kanila na perpektong mag-navigate sa dagat ng panahon ng Cambrian.

Ang mga Chordates ay mga 700 milyong taong gulang, ngunit mabuti kasaysayang heolohikal ang kanilang ebolusyon ay nagsisimula lamang sa Cambrian. Sa mga karagatan ng Cambrian at Ordovician, ang mga naturang chordates lamang ang kilala na walang mga panga.
Sa Silure, nangyari, nangyari, nangyari, nabuo ang mga amphibian na may mga panga ... Ang mga pating at sinag, isda ng sturgeon, bony fish at amphibian ay nanggaling sa kanila.
Sa panahon ng Carboniferous, lumitaw ang mga terrestrial na hayop - anopsid ... Ang mga synopsid at diopsid ay nagmula sa mga anopsid. Ang pag-uuri na ito, tulad nito, ay isinasaalang-alang ang bilang ng mga temporal na hukay, ngunit sa katotohanan ay hindi ito ganoon. Halimbawa, ang mga anopsid ankylosaur ay inuri bilang mga diopsid, at ang mga ibon na may bukas na temporal depressions, mga hollows ... Kasama sa mga synopsid ang mga monotreme, na katulad ng istraktura ng bungo sa mga dinosaur at ibon. Ang mga monotreme ay inuri bilang mga mammal dahil sa kanilang mga pagtatago sa balat at lana. Ngunit ang mga pagtatago ng monotremes ay hindi tulad ng gatas, ngunit ang pagtatago ng coccygeal gland ng mga ibon, at ang lana ay kapareho ng buhok ng mga pterosaur ...
Sa pangkalahatan, maraming klase ng mga hayop ang nabuo mula sa mga anopsid, na pinagsasama at nabubuo ang kanilang mga katangian sa iba't ibang paraan. Ang mga putot na ito ay: mga butiki at ahas, mga buwaya, mga dinosaur (kabilang ang mga monotreme), mga plesiosaur, mga theromorph (kabilang ang mga ichthyosaur).
Ang mga inapo ng theromorph, mammal, ay kilala sa Triassic at Cretaceous. Ngunit ang mga mammal ay nakatanggap ng pagkakataon na kumalat nang malawakan lamang sa panahon ng Paleogene at Neogene. Naging posible ito pagkatapos ng malawakang pagkalipol sa Upper Cretaceous ng maraming species sa lahat ng klase ng vertebrates.
Sa Paleogene, lumitaw ang mga mammal na may nakatiklop na cerebral cortex: predatory, hoofed-hyrax, monkeys. Ang ninuno ng lahat ng mga order na ito ng mga mammal ay ilang uri ng mga lemur. Ang lahat ng mga sinaunang mammal na may nakatiklop na cerebral cortex ay katulad ng mga lemur.
Nawala ang lahat ng buto ng mga pating sa proseso ng ebolusyon
29.05.2015 14:22
Ang isang fossil shark na natagpuan sa kanlurang Australia ay literal na nagpabaligtad ng mga ideya ng mga siyentipiko tungkol sa ebolusyon ng grupong ito ng isda. Kung hanggang kamakailan lamang ay itinuturing na mga primitive na nilalang ang mga mapanganib na mandaragit sa dagat, ngayon ay kailangang ituring sila ng mga biologist bilang napaka-advanced na mga nilalang na dumating sa isang mahabang ebolusyonaryong landas.

Gogoselachus lynnbeazleyae. Muling pagtatayo: John Long
Ang rebolusyon sa ichthyology ay itinanghal ng paleontologist na si John Long ng Flinders University. Matagal nang gumugol ng tatlong dekada sa paghuhukay ng Devonian Gogo formation sa rehiyon ng Kimberley. Noong 2005, natagpuan niya doon ang fossilized skeleton ng isang pating na nanirahan sa isang mainit na tropikal na dagat 380 milyong taon na ang nakalilipas. Ang pag-aaral at paglalarawan ng hayop ay naantala, at ang artikulo tungkol sa Gogoselachus lynnbeazleyae ay nai-publish ngayon lamang.
"Ang mga pating ay itinuturing na primitive na isda sa batayan na ang kanilang balangkas ay gawa sa kartilago at hindi sila bumubuo ng mga buto," sabi ni Propesor Long. "Gayunpaman, ibinabalik natin ngayon ang ideyang ito, na pinagtatalunan na ang mga naunang fossil shark ay talagang may tunay na kalansay ng buto, at pagkatapos ay nawala lamang ito."
Talaga, tissue ng buto sa modernong mga pating, ito ay matatagpuan lamang sa mga ugat ng ngipin, at ang balangkas at maging ang bungo ay gawa sa cartilaginous tissue, na itinuturing na pasimula ng bone tissue. Ang mga ideyang ito ay pinalawak sa lahat ng mga pating ng nakaraan. Ngunit nang tingnan ni Long ang gogoselachus cartilage sa ilalim ng mataas na pagpapalaki gamit ang microtomography, nakita niya dito ang mga tunay na osteocytes - ang mga selula na bumubuo sa mga buto.
"Ang aming fossil shark ay nagpakita sa unang pagkakataon ng isang tunay na bony structure na nagtatali ng maliliit na cartilage. Kaya't nakakakita kami ng isang pating na talagang nagmula sa isang taong may mas maraming buto sa kanilang balangkas. At sa kabilang dulo ng linyang ito ay ang mga modernong pating na ganap na nawala ang kanilang mga buto at naging cartilaginous. Kaya, ang aming fossil ay nagpapahintulot sa amin na obserbahan ang ebolusyon ng mga tisyu, at ipinaliwanag ang mga dahilan kung bakit ang mga modernong pating ay naging matagumpay ngayon - sila ay nagbigay lamang ng mga buto upang maging mas magaan, "paliwanag ni Long.
"Talaga naman kawili-wiling pagtuklas Sinabi ni Per Ahlberg, propesor ng paleontology sa Uppsala University. – Ang mga kalansay ng mga modernong pating ay binubuo ng isang kakaibang tisyu na tinatawag na prismatic calcified cartilage. Ang cartilage na ito ay mineralized at hindi mukhang matigas na sheet, ngunit tulad ng isang mosaic ng maliliit na mineral prisms. Ang nasabing tissue ay medyo naiiba sa buto, at ang pinagmulan nito ay hindi pa lubos na nauunawaan. Ang bagong pating mula sa Gogo ay nagpapakita na ang unang bersyon ng prismatic calcified cartilage ay tila may mga puwang sa pagitan ng mga prisma na puno ng mga selula ng buto, hindi tulad ng mga modernong.
"Ang pag-aaral ng mga pating ay katulad ng paglutas ng isang malaking palaisipan," dagdag ni Propesor Long. "Sila ay lumitaw 250 milyong taon bago ang mga huling dinosaur at hindi gaanong nagbago mula noon, matagumpay na naabot ang panalong formula. Ngunit kahit na ang kanilang hitsura ay nanatiling halos pareho, ang istraktura ng mga tisyu ay sumailalim sa malalaking pagbabago.
Matagal na pinamamahalaang upang malaman ang lahat ng mga detalyeng ito salamat sa mga tiyak na kondisyon ng pangangalaga ng fossil na materyal sa pagbuo ng Gogo. Karaniwan, ang mga isda ng Devonian ay pinapanatili sa mga fossil na lubos na deformed, na pinatag ng presyon ng mga sedimentary na bato. Ngunit dito sa Kimberley, ang mga sinaunang isda ay dumating sa amin na tatlong-dimensional at napakalaki salamat sa mga carbonate nodules na nabuo sa site ng isang sponge-algae reef.
“Noong panahong iyon, puspusan ang buhay dito, maraming uri ng isda ang nabubuhay, halimbawa, ang mga long-extinct armored placoderms at early bony fish, ang mga inapo nito ay nangingibabaw ngayon. Inaasahan naming makakita ng maraming pating dito, ngunit sa ilang kadahilanan ay hindi sila karaniwan sa bahura na ito,” dagdag ni Long.
Para sa kadahilanang ito, ang pagtuklas ng balangkas ng isang 75-sentimetro na pating ay nakakuha ng maraming pansin. Ang parehong mga sanga ng ibabang panga, mga fragment ng sinturon ng balikat na sumusuporta sa mga palikpik ng pectoral, mga arko ng gill, mga 80 ngipin at ilang daang kaliskis ay nahulog sa mga kamay ng mga siyentipiko. Ang ispesimen ay sumailalim sa maraming iba't ibang mga pag-aaral, at ang isa sa mga ito ay humantong sa isang kahindik-hindik na resulta, na pumipilit sa amin na muling isaalang-alang ang parehong maagang ebolusyon ng mga pating at ang saloobin sa kanila bilang isang primitive, naantalang grupo sa kanilang pag-unlad.
Unang Pating mula sa Late Devonian (Frasnian) Gogo Formation, Nagbigay ng Bagong Liwanag ang Kanlurang Australia sa Pagbuo ng Tessellated Calcified Cartilage na inilathala ng PLOS ONE
Doi: 10.1371/journal.pone.0126066
Ayon sa bagong data mula sa mga Amerikanong siyentipiko, ang mga ibon ay hindi mga inapo ng mga dinosaur at nagmula sa isang espesyal na grupo ng mga archosaur na humiwalay sa mga higanteng butiki sa malayong nakaraan.

Scansoriopteryx. Muling pagtatayo: Matt Martyniuk
Isang kahindik-hindik na pagtuklas na maaaring literal na ibalik ang lahat ng modernong paleontolohiya sa ulo nito ay ginawa ni Stephen Cherkas ng Blanding Dinosaur Museum at Alan Feduccia ng University of North Carolina. Gamit ang kanilang bagong pamamaraan, pinag-aralan nila ang mga labi ng maliliit na balahibo na pangolin Scansoriopteryx at dumating sa konklusyon na walang dahilan upang ituring itong isang dinosaur.
Ang Scansoriopteryx, na ang pangalan ay isinalin sa Russian na tinatayang bilang "wing-flyer" o "lap-wing", ay isang maliit na archosaur, humigit-kumulang sa laki ng isang maya. Ang mga labi ng nag-iisang, wala pang gulang na indibidwal ng Scansoriopteryx ay natagpuan sa simula ng ika-21 siglo sa mga deposito ng Jurassic ng lalawigan ng Liaoning ng Tsina. Sa paghusga sa pamamagitan ng mga fossil na nakaligtas hanggang sa araw na ito, ang Scansoriopteryx ay inayos nang mas primitive kaysa sa sikat na Archaeopteryx at mahusay na nakaakyat sa mga puno, lumilipad mula sa kanila sa tulong ng maliliit na pakpak.
Hanggang kamakailan lamang, ang Scansoriopteryx ay naiugnay sa mga coelurosaur, isang pangkat ng mga theropod kung saan, ayon sa karamihan ng mga siyentipiko, nagmula ang mga modernong ibon. Gayunpaman, ang pag-aaral ng Cherkas (na, sa pamamagitan ng paraan, natuklasan Scansoriopteryx) at Feduccia pinipilit sa amin upang muling isaalang-alang ang diskarteng ito. Gumamit ang duo ng mga siyentipiko ng 3D microscopy at low-angle photography para ipaliwanag ang mga istrukturang hindi pa malinaw na nakikita noon. Salamat dito, nagawang pinuhin ng mga paleontologist ang mga natural na balangkas ng mga buto ng pelvis, buntot at paa, at sa parehong oras ay nakahanap ng mga pinahabang tendon na nakaunat sa kahabaan ng tail vertebrae, tulad ng sa Velociraptor.
Gayunpaman, karamihan sa mga ebidensya ay nagmumungkahi na ang Scansoriopteryx ay kulang sa mga pangunahing tampok na istruktura ng kalansay upang maging kwalipikado ito bilang isang dinosaur. Sa halip, siya ay isang inapo ng mga sinaunang archosaur na dalubhasa sa pag-akyat ng puno bago pa man lumitaw ang mga kakila-kilabot na butiki. magpinsan.
Samantala, ang mga tipikal na avian adaptation ay malinaw na nakikita sa Scansoriopteryx mismo, tulad ng mga pahabang forelimbs na naging feathered wings, isang espesyal na lunate carpal bone at inangkop sa pag-upo sa mga sanga ng paa. Malamang, ang hayop na ito ay nasa simula ng pag-unlad ng paglipad, kung saan ito umalis, na dumausdos mula sa mga sanga ng mga puno.