Sa kauna-unahang pagkakataon, lumilitaw ang respiratory system sa marine annelids - mga sandworm at nereid, kung saan ang mga primitive na hasang ay matatagpuan sa mga sanga ng dorsal ng parapodia. Bilang karagdagan, sa mga annelids, ang palitan ng gas ay nangyayari sa pamamagitan ng balat na mayaman sa mga daluyan ng dugo.
Sa mga mammal at mollusk, ang istraktura ng mga organ ng paghinga ay nakasalalay sa mga kondisyon ng kanilang tirahan: sa mga aquatic form, ito ay mga hasang na maaaring gumamit ng oxygen na natunaw sa tubig; sa terrestrial - baga at trachea, inangkop sa paggamit ng atmospheric oxygen.
Kaya, ang mga crustacean ay may mga hasang, na matatagpuan sa mga binti at panga sa ilalim ng mga lateral folds ng cephalothoracic shield, kung saan sila ay patuloy na hinuhugasan ng tubig.
Ang sistema ng paghinga ng mga arachnid ay kinakatawan ng alinman sa hugis-dahon na baga o tracheae. Parehong bukas palabas na may mga espesyal na pagbubukas - stigmas sa mga lateral na bahagi ng mga segment. Sa lung sac mayroong maraming mga leaf folds kung saan dumadaan ang mga capillary ng dugo. Ang mga baga ng arachnid ay homologous sa hasang ng mga crustacean. Ang trachea ay isang sistema ng mga branched tube na direktang napupunta sa lahat ng mga organo, kung saan nagaganap ang pagpapalitan ng tissue.
Ang mga insekto ay humihinga sa pamamagitan ng trachea.
Sa maraming mga arthropod, na may manipis na chitinous na takip at medyo malaki ang ibabaw ng katawan, ang diffuse respiration ay sinusunod din.
Sa mga mollusc, ang mga pangunahing organ sa paghinga ay mga hasang, maliban sa mga terrestrial mollusc, halimbawa, ang ilang mga gastropod, na nawalan ng mga hasang, at ang kanilang mantle cavity ay naging baga.
Maraming mga invertebrate ang may mga device na nagpapataas ng respiratory surface sa anyo ng mga lokal na dalubhasang respiratory organ.
Ebolusyon ng sistema ng paghinga sa mga chordates .
Ang sistema ng paghinga ng lahat ng chordates, pareho sa pinagmulan at topographically, ay konektado sa endoderm.
Sa aquatic chordates, ang function ng paghinga ay ginagampanan ng gill slits na tumatagos sa anterior section ng intestinal tube-pharynx. Sa terrestrial chordates, ang gill slits ay nagsasara sa panahon ng embryonic development at pagkatapos ay nawawala. Ang pag-andar ng paghinga ay ginagawa ng mga baga, na nabuo mula sa pag-usli ng tubo ng bituka. Ang ebolusyon ng gill apparatus sa chordates ay ipinahayag sa isang pagbawas sa bilang ng mga gill slits na may sabay-sabay na pagtaas sa respiratory surface sa pamamagitan ng pagbuo ng mga gill filament.
Ang ebolusyon ng mga baga ay napunta sa direksyon ng paghihiwalay ng respiratory tract at isang pagtaas sa respiratory surface sa pamamagitan ng pagbuo ng mga baga ng isang spongy na istraktura na may isang kumplikadong sistema ng sumasanga ng intracellular bronchi, na nagtatapos sa mga vesicle na may mga selulang selula.
Ang pinaka-primitive na sistema ng paghinga ay nasa lancelet, na kabilang sa mas mababang chordates (subtype Cranial). Ang nauuna na bituka (pharyngeal wall) ay natatakpan ng mga gill slits (hanggang sa 150 pares), na bumubukas sa arterial (periobranchial) na lukab.
Sa mga cyclostomes (subtype na Vertebrates), ang mga organ ng paghinga ay mga gill slits din, ngunit mas kaunti na ang mga ito (5-15 pares). Nakikipag-usap sila sa anterior na bituka at bukas sa labas na may mga independiyenteng pagbubukas.
Ang mga tunay na hasang ay lumilitaw sa mga chordates sa isda. Ang mga ito ay mga manipis na fold ng mauhog lamad ng pharynx, na nakahiga sa mga arko ng hasang at binibigyan ng venous blood sa pamamagitan ng branchial arteries, na bumubuwag dito sa mga capillary. Ang isda ay may 4-7 (karaniwan ay 5) gill sac sa pagitan ng mga arko ng hasang. Ang mga stamen na matatagpuan sa matambok na ibabaw ng mga arko ng hasang ay pumipigil sa pagpasok ng pagkain sa mga hasang mula sa pharynx.
Bilang karagdagan sa mga hasang, ang mga isda ay may karagdagang mga organ sa paghinga na nagpapahintulot sa kanila na gumamit ng atmospheric oxygen. Ang nasabing organ sa isda ay ang swim bladder. Ang mga dingding nito ay mayaman sa mga daluyan ng dugo, samakatuwid, sa ilang mga isda na lumulubog sa silt, maaari itong magsilbi para sa palitan ng gas. Ang swim bladder sa karamihan ng mga isda ay nabubuo mula sa dorsal na bahagi ng pharynx at hindi isang homologue ng mga baga. Tanging sa mga isda na may lobe-finned ang swim bladder ay nabuo bilang isang protrusion ng ventral na bahagi ng pharynx at nagsisilbing homologue ng mga baga ng mga terrestrial na hayop, dahil ang mga baga ng vertebrates ay bubuo mula sa ventral na bahagi ng gill sac.
Ang lungfish ay may mga baga ngunit walang swim bladder.
Sa amphibian larvae, tulad ng sa isda, ang mga organ ng paghinga ay kinakatawan ng mga panlabas na hasang na sumasanga ng puno. Sa karamihan ng mga amphibian na nasa hustong gulang, ang mga baga ay lumilitaw sa anyo ng manipis na pader na magkapares na mga outgrowth ng ventral pharynx sa likod ng huling gill sac. Dahil sa kawalan ng dibdib at dayapragm, ang hangin ay pumapasok sa kanila mula sa oral cavity dahil sa mga paggalaw ng paglunok na isinasagawa ng chin-hyoid na kalamnan. Ang materyal ng mga arko ng hasang kasunod ng arko ng hypoglossal ay bahagi ng kartilago ng larynx, na lumilitaw sa unang pagkakataon sa mga amphibian, bilang ang unang organ na nauugnay sa mas mababang respiratory tract. Ang mga baga ay nagsisimula nang direkta mula sa larynx. Ang mga ito ay coarse-meshed at may maliit na respiratory surface, at samakatuwid, gas exchange in higit pa Ito ay isinasagawa sa pamamagitan ng balat, na kung saan ay natagos ng isang malaking bilang ng mga capillary ng dugo at ibinibigay ng mga mucous glandula.
Sa mga reptilya, ang sistema ng paghinga ay kumplikado. Ang kanilang mga baga ay pinong cellular na, naglalaman sila ng maraming cellular crossbars at may malaking respiratory surface. Sa respiratory tract, na may kaugnayan sa pangwakas na paglabas ng mga reptilya sa lupa, ang pag-unlad ay sinusunod: ang itaas na respiratory tract ay nakikilala, kahit na hindi ganap na natanggal mula sa oral cavity - ito ang ilong na lukab, at ang mas mababang mga ay ang larynx, trachea at bronchi. Sa unang pagkakataon, lumilitaw ang dayapragm, na kumukuha ng passive na bahagi sa paghinga. Bahagyang pinaghihiwalay nito ang dibdib at lukab ng tiyan, o wala ang mga hibla ng kalamnan.
Ang mekanismo ng paghinga ay isinasagawa sa pamamagitan ng pag-urong ng mga intercostal na kalamnan, na nagtatakda ng paggalaw ng dibdib.
Sa mga ibon, ang mga baga ay makapal na spongy na katawan na tinusok ng mga ramification ng bronchi, at hindi mga bag, tulad ng sa mga reptilya. Bilang karagdagan, ang mga ito ay pupunan bilang isang air reservoir na may manipis na pader na air bag. Ang huli ay matatagpuan sa pagitan ng lahat ng mga organo ng hangin, sa pagitan ng mga kalamnan, sa lukab ng buto at sa ilalim ng buto.
Kapag ang mga pakpak ay nakataas, ang mga air sac sa pamamagitan ng mga baga ay puno ng hangin, kapag ang mga pakpak ay ibinaba, ang hangin ay inilabas sa pamamagitan ng mga baga. Kaya, sa panahon ng paglipad, ang mga ibon ay nagsasagawa ng dobleng paghinga. Sa panahon ng pahinga, ang ibon ay humihinga lamang sa pamamagitan ng pagpapalawak at pagkontrata ng dibdib.
Ang mga organ ng paghinga ng lahat ng mga mammal ay nailalarawan sa pagiging kumplikado ng parehong mga baga, na may istraktura ng alveolar, at ang respiratory tract. Mayroong karagdagang sangay ng bronchial tree kung ihahambing sa mga nakaraang klase ng vertebrates. Ang trachea ay nahahati sa bronchi, na nagsanga sa bronchi ng ikalawa, ikatlo at ikaapat na mga order at sa pinakamaliit mismo - mga bronchioles, sa mga sanga kung saan mayroong alveoli, mga pulmonary vesicle na may malaking lugar sa bag (mga 90 cm squared), kung saan nagaganap ang palitan ng gas. Ang pangunahing kalamnan na naglalaro mahalagang papel sa gawa ng paghinga, ay ang dayapragm. Ang respiratory tract ng mga mammal ay may linya na may ciliated epithelium at ganap na nakahiwalay sa digestive system.
Ang mga organ ng paghinga ng mga vertebrates ay may dalawang uri - gills at baga, at sa isang makabuluhang bahagi ng vertebrates, ang balat ay mahalaga sa paghinga. Ang gill apparatus ay isang sistema ng magkapares, kadalasang simetriko ang kinalalagyan, slits na nagsisilbing komunikasyon ng pharynx sa panlabas na kapaligiran. Ang anterior at posterior wall ng gill slits ay may linya na may mucous membrane na bumubuo ng lamellar outgrowths; ang mga outgrowth ay nahahati sa mga petals, na tinatawag na hasang. Ang bawat gill plate sa itaas ng mga petals ay tinatawag na half-gill. Sa pagitan ng mga gill slits (sa gill septa) ay ang visceral gill arches. Kaya, ang bawat gill arch ay konektado sa dalawang kalahating hasang ng dalawang magkaibang hasang slits. Ang mga organ sa paghinga ng mga terrestrial vertebrates - ang mga baga - sa diagram ay isang pares ng mga sac na bumubukas sa pharynx sa pamamagitan ng laryngeal fissure. Sa embryonally, ang mga baga ay bumangon sa anyo ng isang protrusion ng dingding ng tiyan ng pharynx sa likod ng gill apparatus, iyon ay, sila ay mula sa endodermal na pinagmulan. Sa mga unang yugto ng pag-unlad ng embryonic, ang mga buds ng baga ay kahawig ng isang pares ng panloob (endodermal) gill slits. Ang mga pangyayaring ito, pati na ang suplay ng dugo at mga tampok ng innervation na karaniwan sa mga baga at hasang, ay nagpipilit sa amin na isaalang-alang ang mga baga bilang mga homologue ng posterior na pares ng mga gill sac. Ang balat ay kasangkot sa paghinga sa mga kaso kung saan walang siksik na sungay o bony na kaliskis dito, halimbawa, sa mga amphibian, hubad na balat na isda. Sa paggana, ang sistema ng paghinga ay kasangkot sa pagpapayaman ng dugo na may oxygen at sa pag-alis ng carbon dioxide. Ang ammonia ay inilalabas sa pamamagitan ng respiratory system sa mas mababang mga hayop sa tubig. Sa mga hayop na may mainit na dugo, ito ay kasangkot sa mga proseso ng thermoregulation. Ang prinsipyo ng pagpapatakbo ng sistema ng paghinga ay ang pagpapalitan ng CO2 at O2 sa pagitan ng mga daloy ng gas at dugo, na nakadirekta sa countercurrent patungo sa isa't isa. Ang paghinga ay isang proseso na kumukuha ng oxygen at naglalabas ng carbon dioxide. Ang sistema ng paghinga ay gumaganap ng pinakamahalagang pag-andar - gas exchange, kung wala ang buhay ay imposible, dahil ang pagbabagong-anyo ng enerhiya sa katawan ay nangyayari bilang isang resulta ng oxidative decay sustansya na may partisipasyon ng oxygen. palitan ng gas sa baga. Ang dugo na dumadaloy sa mga baga mula sa puso (venous) ay naglalaman ng kaunting oxygen at maraming carbon dioxide; ang hangin sa alveoli, sa kabaligtaran, ay naglalaman ng maraming oxygen at mas kaunting carbon dioxide. Bilang resulta, ang two-way diffusion ay nangyayari sa pamamagitan ng mga dingding ng alveoli at mga capillary. pumapasok ang oxygen sa dugo, at ang carbon dioxide ay gumagalaw mula sa dugo patungo sa alveoli. Sa dugo, ang oxygen ay pumapasok sa mga pulang selula ng dugo at pinagsama sa hemoglobin. Ang oxygenated na dugo ay nagiging arterial at pumapasok sa kaliwang atrium sa pamamagitan ng mga pulmonary veins. pagpapalitan ng gas sa mga tisyu. Ang pagpapalitan ng mga gas sa mga tisyu ay isinasagawa sa mga capillary. Sa pamamagitan ng kanilang manipis na mga pader, ang oxygen ay pumapasok mula sa dugo papunta sa tissue fluid at pagkatapos ay sa mga cell, at ang carbon dioxide mula sa mga tissue ay pumapasok sa dugo. Ang konsentrasyon ng oxygen sa dugo ay mas malaki kaysa sa mga selula, kaya madali itong kumalat sa kanila. Ang konsentrasyon ng carbon dioxide sa mga tisyu kung saan ito nakolekta ay mas mataas kaysa sa dugo. Samakatuwid, ito ay pumasa sa dugo, kung saan ito nagbubuklod mga kemikal na compound plasma at bahagyang may hemoglobin, ay dinadala ng dugo sa mga baga at inilabas sa atmospera.
5.1. Pinagmulan at pag-andar ng sistema ng paghinga.
Ang sistema ng paghinga ng mga organismo ng hayop ay mula sa endodermal na pinagmulan, dahil sa pinagmulan ito ay nauugnay sa sistema ng pagtunaw. Ang pag-andar ng mga organ sa paghinga ay ang pagpapalitan ng gas sa pagitan ng isang partikular na organismo at ng kapaligiran.
5.2. Ang ebolusyon ng respiratory system sa isang bilang ng mga invertebrates.
Sa kauna-unahang pagkakataon, lumilitaw ang respiratory system sa marine annelids - mga sandworm at nereid, kung saan ang mga primitive na hasang ay matatagpuan sa mga sanga ng dorsal ng parapodia. Bilang karagdagan, sa mga annelids, ang palitan ng gas ay nangyayari sa pamamagitan ng balat na mayaman sa mga daluyan ng dugo.
Sa mga mammal at mollusk, ang istraktura ng mga organ ng paghinga ay nakasalalay sa mga kondisyon ng kanilang tirahan: sa mga aquatic form, ito ay mga hasang na maaaring gumamit ng oxygen na natunaw sa tubig; sa terrestrial - baga at trachea, inangkop sa paggamit ng atmospheric oxygen.
Kaya ang mga crustacean ay may mga hasang, na matatagpuan sa mga binti at panga sa ilalim ng mga lateral folds ng cephalothoracic shield, kung saan sila ay patuloy na hinuhugasan ng tubig.
Ang sistema ng paghinga ng mga arachnid ay kinakatawan ng alinman sa hugis-dahon na baga o tracheae. Parehong bukas palabas na may mga espesyal na pagbubukas - stigmas sa mga lateral na bahagi ng mga segment. Sa lung sac mayroong maraming mga leaf folds kung saan dumadaan ang mga capillary ng dugo. Ang mga baga ng arachnid ay homologous sa hasang ng mga crustacean. Ang trachea ay isang sistema ng mga branched tube na direktang napupunta sa lahat ng mga organo, kung saan nagaganap ang pagpapalitan ng tissue.
Ang mga insekto ay humihinga sa pamamagitan ng trachea.
Sa maraming mga arthropod, na may manipis na chitinous na takip at medyo malaki ang ibabaw ng katawan, ang diffuse respiration ay sinusunod din.
Sa mga mollusc, ang mga pangunahing organ sa paghinga ay mga hasang, maliban sa mga terrestrial mollusc, halimbawa, ang ilang mga gastropod, na nawalan ng mga hasang, at ang kanilang mantle cavity ay naging baga.
Maraming mga invertebrate ang may mga device na nagpapataas ng respiratory surface sa anyo ng mga lokal na dalubhasang respiratory organ.
5.3. Ang ebolusyon ng respiratory system sa chordates.
Ang sistema ng paghinga ng lahat ng chordates, kapwa sa pinagmulan at topographically, ay konektado sa bituka, iyon ay, sa endoderm.
Sa aquatic chordates, ang function ng paghinga ay ginagampanan ng gill slits na tumatagos sa anterior section ng intestinal tube-pharynx. Sa terrestrial chordates, ang gill slits ay nagsasara sa panahon ng embryonic development at pagkatapos ay nawawala. Ang pag-andar ng paghinga ay ginagawa ng mga baga, na nabuo mula sa pag-usli ng tubo ng bituka.
Ang ebolusyon ng gill apparatus sa chordates ay ipinahayag sa isang pagbawas sa bilang ng mga gill slits na may sabay-sabay na pagtaas sa respiratory surface sa pamamagitan ng pagbuo ng mga gill filament.
Ang ebolusyon ng mga baga ay napunta sa direksyon ng paghihiwalay ng respiratory tract at isang pagtaas sa respiratory surface sa pamamagitan ng pagbuo ng mga baga ng isang spongy na istraktura na may isang kumplikadong sistema ng sumasanga ng intracellular bronchi, na nagtatapos sa mga vesicle na may mga selulang selula.
Ang pinaka-primitive na sistema ng paghinga ay nasa lancelet, na kabilang sa mas mababang chordates (subtype Cranial). Ang nauuna na seksyon ng bituka (pharyngeal wall) ay butas-butas ng gill slits (hanggang sa 150 pares), na bumubukas sa arterial (periobranchial) na lukab.
Sa mga cyclostomes (subtype na Vertebrates), ang mga organ ng paghinga ay mga gill slits din, ngunit mas kaunti na ang mga ito (5-15 pares). Nakikipag-usap sila sa anterior na bituka at bukas sa labas na may mga independiyenteng pagbubukas.
Ang mga tunay na hasang ay lumilitaw sa mga chordates sa isda. Ang mga ito ay mga manipis na fold ng mauhog lamad ng pharynx, na nakahiga sa mga arko ng hasang at binibigyan ng venous blood sa pamamagitan ng branchial arteries, na bumubuwag dito sa mga capillary. Ang isda ay may 4-7 (karaniwan ay 5) gill sac sa pagitan ng mga arko ng hasang. Ang mga stamen na matatagpuan sa matambok na ibabaw ng mga arko ng hasang ay pumipigil sa pagpasok ng pagkain sa mga hasang mula sa pharynx.
Bilang karagdagan sa mga hasang, ang mga isda ay may karagdagang mga organ sa paghinga na nagpapahintulot sa kanila na gumamit ng atmospheric oxygen. Ang nasabing organ sa isda ay ang swim bladder. Ang mga dingding nito ay mayaman sa mga daluyan ng dugo, samakatuwid, sa ilang mga isda na lumulubog sa silt, maaari itong magsilbi para sa palitan ng gas. Ang swim bladder sa karamihan ng mga isda ay nabubuo mula sa dorsal na bahagi ng pharynx at hindi isang homologue ng mga baga. Tanging sa mga isda na may lobe-finned ang swim bladder ay nabuo bilang isang protrusion ng ventral na bahagi ng pharynx at nagsisilbing homologue ng mga baga ng mga terrestrial na hayop, dahil ang mga baga ng vertebrates ay bubuo mula sa ventral na bahagi ng gill sac.
Ang lungfish ay may mga baga ngunit walang swim bladder.
Sa amphibian larvae, tulad ng sa isda, ang mga organ ng paghinga ay kinakatawan ng mga panlabas na hasang na sumasanga ng puno. Sa karamihan ng mga amphibian na nasa hustong gulang, ang mga baga ay lumilitaw sa anyo ng manipis na pader na magkapares na mga outgrowth ng ventral pharynx sa likod ng huling gill sac. Dahil sa kawalan ng dibdib at dayapragm, ang hangin ay pumapasok sa kanila mula sa oral cavity dahil sa mga paggalaw ng paglunok na isinasagawa ng chin-hyoid na kalamnan. Ang materyal ng mga arko ng hasang kasunod ng arko ng hypoglossal ay bahagi ng kartilago ng larynx, na lumilitaw sa unang pagkakataon sa mga amphibian, bilang ang unang organ na nauugnay sa mas mababang respiratory tract. Ang mga baga ay nagsisimula nang direkta mula sa larynx. Ang mga ito ay coarse-meshed at may maliit na respiratory surface, at samakatuwid ang gas exchange ay isinasagawa sa mas malawak na lawak sa pamamagitan ng balat, na kung saan ay natagos ng isang malaking bilang ng mga capillary ng dugo at ibinibigay sa mga mucous glands.
Sa mga reptilya, ang sistema ng paghinga ay kumplikado. Ang kanilang mga baga ay pinong cellular na, naglalaman sila ng maraming cellular crossbars at may malaking respiratory surface. Sa respiratory tract, na may kaugnayan sa pangwakas na paglabas ng mga reptilya sa lupa, ang pag-unlad ay sinusunod: ang itaas na respiratory tract ay nakikilala, kahit na hindi ganap na natanggal mula sa oral cavity - ito ang ilong na lukab, at ang mas mababang mga ay ang larynx, trachea at bronchi. Sa unang pagkakataon, lumilitaw ang dayapragm, na kumukuha ng passive na bahagi sa paghinga. Bahagyang pinaghihiwalay nito ang dibdib at lukab ng tiyan, o wala ang mga hibla ng kalamnan.
Ang mekanismo ng paghinga ay isinasagawa sa pamamagitan ng pag-urong ng mga intercostal na kalamnan, na nagtatakda ng paggalaw ng dibdib.
Sa mga ibon, ang mga baga ay makapal na spongy na katawan na tinusok ng mga ramification ng bronchi, at hindi mga bag, tulad ng sa mga reptilya. Bilang karagdagan, ang mga ito ay pupunan bilang isang air reservoir na may manipis na pader na air bag. Ang huli ay matatagpuan sa pagitan ng lahat ng mga organo ng hangin, sa pagitan ng mga kalamnan, sa lukab ng buto at sa ilalim ng buto.
Kapag ang mga pakpak ay nakataas, ang mga air sac sa pamamagitan ng mga baga ay puno ng hangin, kapag ang mga pakpak ay ibinaba, ang hangin ay inilabas sa pamamagitan ng mga baga. Kaya, sa panahon ng paglipad, ang mga ibon ay nagsasagawa ng dobleng paghinga. Sa panahon ng pahinga, ang ibon ay humihinga lamang sa pamamagitan ng pagpapalawak at pagkontrata ng dibdib.
Ang mga organ ng paghinga ng lahat ng mga mammal ay nailalarawan sa pagiging kumplikado ng parehong mga baga, na may istraktura ng alveolar, at ang respiratory tract. Mayroong karagdagang sangay ng bronchial tree kung ihahambing sa mga nakaraang klase ng vertebrates. Ang trachea ay nahahati sa bronchi, na nagsanga sa bronchi ng ikalawa, ikatlo at ikaapat na mga order at sa pinakamaliit mismo - mga bronchioles, sa mga sanga kung saan mayroong alveoli, mga pulmonary vesicle na may malaking lugar sa bag (mga 90 cm squared), kung saan nagaganap ang palitan ng gas. Ang pangunahing kalamnan na gumaganap ng mahalagang papel sa pagkilos ng paghinga ay ang dayapragm. Ang respiratory tract ng mga mammal ay may linya na may ciliated epithelium at ganap na nakahiwalay sa digestive system.
5.4. Anomalya at malformations ng respiratory system sa mga tao.
1. Sa embryogenesis ng tao, ang unang koneksyon sa pagitan ng digestive at respiratory system ay makikita. Kaugnay nito, nagiging malinaw na ang isang tao ay may isang malaking grupo ng mga congenital malformations ng esophagus at trachea ng uri esophagotracheal fistula (« mga biyak ng hasang»).
2. Dysontogenetic bronchopulmonary cyst- isang bilugan na guhit sa baga, na nililimitahan mula sa nakapaligid na tisyu sa pamamagitan ng isang primitively constructed na pader ng isang hindi nakikilalang bronchus. Ang mga anomalyang ito ay maaaring ipaliwanag sa pamamagitan ng isang paglabag sa alveolar differentiation ng tissue ng baga.
3. Cystic hypoplasia- underdevelopment ng baga. Sa patolohiya na ito, ang buong lobes ng baga ay kulang sa pag-unlad, na maraming mga cavity na nauugnay sa malaking bronchi at pagkakaroon ng isang maliit na ibabaw. Ang palitan ng gas sa naturang mga lugar ng baga ay nanghina nang husto.
4. Hypoplasia ng diaphragm- hindi pag-unlad ng diaphragm mula sa maliliit na depekto sa simboryo nito upang makumpleto ang aplasia. Ang anomalyang ito ay hindi tugma sa buhay at nangyayari nang mas madalas kasama ng iba pang maraming malformation.
Lektura 12
Mga Class Amphibian (Amphibia)
Katangian ng klase
Ang mga amphibian ay ang mga unang vertebrate na dumapo sa lupa, ngunit hindi nawalan ng ugnayan kapaligirang pantubig. Ang pagpaparami ay nangyayari sa tubig, mayroong isang aquatic fish-like larva. Bilang karagdagan, ang isang malapit na kaugnayan sa tubig ay katangian ng karamihan sa mga species ng amphibian kahit na sa pagtanda, kaya mayroon silang mga adaptasyon para sa buhay kapwa sa tubig at sa lupa.
mga pabalat kinakatawan ng balat, malambot, hubad, permeable sa mga gas at tubig.
Skeleton at kalamnan . Ang balangkas ay sumasailalim sa isang bilang ng mga pagbabagong nauugnay sa daan sa lupa buhay. Ang mga servikal at sacral na seksyon ng gulugod ay pinaghihiwalay, na mayroong isang vertebra bawat isa. Ang bungo, sa tulong ng dalawang condyles, ay gumagalaw na nakakabit sa cervical vertebra. Mula sa magkapares na palikpik ng freshwater lobe-finned fish para sa paggalaw sa lupa, ang mga paa ay nabuo, na isang sistema ng mga lever. Ang musculature ay nawawala ang metameric na istraktura nito at kinakatawan ng maraming indibidwal na kalamnan.
Sistema ng paghinga . Ang sistema ng paghinga ng mga amphibian ay kawili-wili dahil sa proseso ng pag-unlad mayroong isang paglipat mula sa paghinga ng gill hanggang sa paghinga ng baga. Ang paghinga ng balat ay napakahalaga. Ang respiratory tract ay hindi maganda ang pag-unlad.
Daluyan ng dugo sa katawan . Dahil lumitaw ang mga baga, nangyayari ang isang pulmonary (maliit) na sirkulasyon. Ang puso ng amphibian ay nagiging tatlong silid, tatlong pares ng mga arko ng arterial ang umaalis dito. Ang metabolismo ay hindi pa masyadong matindi, ang mga amphibian ay hindi mapanatili ang isang pare-parehong temperatura ng katawan, sila ay kabilang sa mga poikilothermic na hayop.
Sistema ng nerbiyos . Sa utak, ang malalaking hemispheres ng forebrain ay pinalaki at ganap na pinaghihiwalay. Ang midbrain at cerebellum ay bahagyang nabuo. 10 pares ng cranial nerves ang umaalis sa utak.
Ang buhay sa himpapawid ay humantong sa paglitaw ng isang bilang ng mga tampok sa mga pandama. Dahil sa convex cornea at flattened lens accommodation ay nagpapabuti. Upang maprotektahan ang mga mata mula sa pagbara at pagkatuyo, ang mga amphibian ay may mga movable eyelids at nictitating membranes. Upang maramdaman ang mga tunog sa hangin, lumilitaw ang tympanic membrane, sa likod nito ay ang air cavity ng gitnang tainga at isang pandinig na buto - ang stirrup, na nagsasagawa ng mga vibrations sa panloob na tainga. Ang Eustachian tube ay nag-uugnay sa gitnang tainga na lukab sa bibig. Lumilitaw ang Choanae, ang mga panloob na butas ng ilong, ang mga daanan ng ilong ay dumaan.
Manifold . Sa modernong fauna, mayroong higit sa 4,000 species ng amphibian, na nahahati sa mga order na Tailless (Anura), Tailed (Caudata) at Legless (Apoda).
Ang mga sumusunod na aromorphoses ay nag-ambag sa paglitaw ng mga amphibian sa lupa: 1. Lumitaw ang liwanag at pulmonary respiration. 2. Pinahirapan ito daluyan ng dugo sa katawan, ang pulmonary circle ng sirkulasyon ng dugo ay nabuo, iyon ay, ang mga amphibian ay may dalawang bilog ng sirkulasyon ng dugo - malaki at maliit. Ang puso ay naging tatlong silid. 4. Nabuo ang magkapares, limang daliri na paa, na isang sistema ng mga lever na may articulated joints at idinisenyo para sa paggalaw sa lupa. 5. Ang isang servikal na rehiyon ay lumitaw sa gulugod, na tiniyak ang paggalaw ng ulo, at isang sacral na rehiyon, ang lugar ng attachment ng pelvic girdle. 6. Lumitaw ang gitnang tainga, talukap ng mata, choanae.
Istruktura at buhay . Ang mga tailed amphibian ay may pahabang katawan, mahabang buntot, at may mga paa. Ang walang buntot ay may isang maikling katawan na pipi sa direksyon ng dorsal-tiyan, ang buntot ay wala. Ang Legless ay may pinahabang katawan na parang bulate na may maraming annular folds ng hubad na balat, walang paa, at maikling buntot.
Ang integument ay kinakatawan ng isang multi-layered epidermis at dermis. Ang balat ay malambot, hubad, natatagusan ng mga gas at tubig, at natagos ng isang network ng mga daluyan ng dugo. Gumagana ito sa mga amphibian bilang isang karagdagang organ sa paghinga, kaya naglalaman ito ng maraming mga glandula na patuloy na naglalabas ng uhog. Ang moisturizing ng balat ay dahil sa ang katunayan na ang pagpapalitan ng gas sa pagitan ng katawan at kapaligiran ay maaaring mangyari lamang sa pamamagitan ng water film. Bilang karagdagan sa mga mucous membrane, mayroon ding mga lason na glandula.
Phylogeny . Devonian Panahon ng Paleozoic, kung saan lumitaw ang mga unang amphibian, tila, ay nailalarawan sa mga pana-panahong tagtuyot, kung saan ang buhay sa maraming mga sariwang tubig ay mahirap para sa mga isda. Ang pag-ubos ng tubig na may oxygen at ang kahirapan sa paglangoy dito ay pinadali ng mga nabubulok na labi ng masaganang mga halaman na tumubo sa panahon ng Carboniferous sa mga latian at sa mga pampang ng mga reservoir. Sa ilalim ng mga kondisyong ito, maaaring tumaas ang ilang isda sa ibabaw at lumunok ng hangin, nabuo ang mga bag sa baga. Ngunit sa matinding pagkatuyo ng mga reservoir, naging imposible na ang buhay para sa mga isda. Hindi makagalaw sa lupa, sila ay namatay. Tanging ang mga aquatic vertebrates, na, kasama ng kakayahan para sa pulmonary respiration, ay nakakuha ng mga limbs na maaaring magbigay ng paggalaw sa lupa, ang makakaligtas sa mga kondisyong ito. Gumapang sila palabas sa lupa at tumawid sa mga kalapit na imbakan ng tubig, kung saan napanatili pa rin ang tubig.
Ang paglipat ng mga vertebrates mula sa isang nabubuhay sa tubig patungo sa isang terrestrial na paraan ng pamumuhay at ang hitsura ng mga amphibian ay sinamahan ng paglitaw ng dalawang mapagpasyang adaptasyon: paghinga ng oxygen sa atmospera at paglipat kasama ang isang solidong substrate. Sa madaling salita, ang paghinga ng hasang ay dapat na pinalitan ng paghinga sa baga, at ang mga palikpik ng mga paa na may limang daliri, na isang sistema ng mga lever na nagsisilbing sumusuporta sa katawan at gumagalaw. Kasabay nito, nagbago din ang ibang mga organ system. Karamihan sa mga mananaliksik ay naniniwala na ang mga amphibian ay nagmula sa freshwater brushfins. ripidistian isda 370-350 milyong taon na ang nakalilipas. Unang lumitaw ichthyostegidae, pagkatapos stegocephalians- shell-headed. Ang mga unang amphibian na lumitaw sa sariwang tubig sa dulo ng Devonian ay ichthyostegids. Ang mga ito ay tunay na transitional form sa pagitan ng lobe-finned fish at amphibian. Kaya, mayroon silang mga simula ng takip ng hasang, isang tunay na buntot ng isda, ang balat ay natatakpan ng maliliit na kaliskis ng isda. Gayunpaman, kasama nito, pinagpares nila ang limang daliri na mga limbs ng terrestrial vertebrates. Ang mga Ichthyostegid ay nabubuhay hindi lamang sa tubig, kundi pati na rin sa lupa. Sa pangkalahatan, ang lahat ng Paleozoic amphibian ay nawala sa panahon ng Triassic.
Mga tampok ng istraktura at pagpaparami.
Kilalanin natin ang mga tampok ng istraktura gamit ang halimbawa ng isang palaka. Ang katawan ay pipi sa direksyon ng dorsal-tiyan, na binubuo ng isang ulo, puno ng kahoy at mga paa. Ang hubad na balat ay basa-basa, hyperosmotic, tubig at bahagi ng oxygen ang pumapasok sa katawan ng palaka sa pamamagitan ng balat. Sa ulo ay nakaumbok na mga mata, na may kaugnayan sa pamumuhay sa hangin, lumitaw ang mga eyelid at isang nictitating membrane. Sa dulo ng muzzle ay ang mga butas ng ilong, ang mga daanan ng ilong ay bumubukas na may mga choan sa oropharyngeal cavity. Sa likod ng mga pagbubukas ng tainga sa mga lalaki ng maraming mga species mayroong mga nagpapalaki na mga bula - mga resonator, sa tulong kung saan ang mga ibinubuga na tunog ay pinalakas. Ang cervical region ay mahina na ipinahayag, ang buntot ay wala. Lever-type limbs, hind five-fingered limbs ay mas mahaba, front four-fingered. Ang pangkulay ay tumatangkilik.
Skeleton at kalamnan . Ang balangkas ng isang palaka, tulad ng lahat ng vertebrates, ay nahahati sa apat na seksyon: ang axial skeleton, ang skeleton ng bungo, ang skeleton ng limbs at ang skeleton ng limb belts.
Ang axial skeleton ay kinakatawan ng gulugod, kung saan, bilang karagdagan sa mga seksyon ng puno ng kahoy at buntot na katangian ng isda, lumitaw ang mga seksyon ng cervical at sacral. Ang bungo ng isang palaka ay movably articulated na may isang solong cervical vertebra sa tulong ng dalawang condyles, na nagsisiguro sa paggalaw ng ulo sa isang vertical na eroplano (ang ulo ay hindi maaaring ilipat sa isang pahalang na eroplano).
Ang bilang ng mga vertebrae ng rehiyon ng puno ng kahoy ay maaaring magkakaiba, ang pinakamalaking bilang ay nasa mga walang paa na amphibian (higit sa isang daan). Ang Anuran amphibian ay may pinakamakaunting vertebrae sa trunk region; ang palaka ay may pito sa kanila. Ang palaka ay walang mga buto-buto, ngunit sa mga buntot na amphibian, ang mga maikling itaas na tadyang ay bubuo sa vertebrae ng puno, at ang mga walang paa ay may tunay na mga buto-buto. Kasama sa rehiyon ng sacral ang isang vertebra, na nagdadala ng mahabang transverse na proseso, kung saan nakakabit ang iliac bones ng pelvis. Ang seksyon ng buntot ng palaka ay nagtatapos sa isang buto ng buntot - urostyle - isang buto, na kung saan ay ilang mga vertebrae na pinagsama sa proseso ng pag-unlad ng embryonic. Kaugnay ng landfall, may mga pagbabago sa bungo, pati na rin ang pagbawas sa mga hasang at gill apparatus.
kanin. 146. Balangkas ng Palaka:
1 - cervical vertebra; 2 - sacral vertebra; 3 - urostyle; 4 - sternum; 5 - buto ng uwak (coracoid); 6 - clavicle; 7 - scapula; 8 - ischium; 9 - ilium; 10 - pubic cartilage; 11 - humerus; 12 - bisig (radius at ulna); 13 - brush; 14 - femur; 15 - mga buto ng ibabang binti (tibia at tibia); 16 - tarsus; 17 - metatarsus; 18 - phalanges ng mga daliri.
Dahil ang tiyak na bigat ng katawan sa lupa ay higit na mas malaki kaysa sa tubig, ang mga amphibian ay may mga paa na nahahati sa mga movably konektado na mga seksyon. Ang ganitong istraktura ay makabuluhang binabawasan ang pagsisikap na ginugol upang mapanatili at ilipat ang katawan sa mga kondisyon ng terrestrial. Ang forelimbs, kadalasang apat na daliri (ang unang daliri ay nabawasan), ay binubuo ng tatlong mga seksyon: ang balikat - ang humerus, ang forearm - ang pinagsamang radius at ulna, at ang kamay, na kinakatawan ng mga buto ng pulso, metacarpus at phalanges ng mga daliri.
Ang mga hind limbs ay binubuo ng tatlong seksyon: hita, ibabang binti at paa. Ang hita ay binubuo ng femur, ang ibabang binti ay gawa sa fused tibia at tibia, ang paa ay gawa sa mga buto ng tarsus, metatarsus at phalanges ng mga daliri.
Ang sinturon sa balikat (Larawan 145) ng palaka ay pumapalibot sa katawan na may malawak na kalahating bilog at nakapirmi sa mga kalamnan. Ito ay kinakatawan ng ilang magkapares na buto: mga talim ng balikat na nagtatapos sa sh

kanin. 145. Sinturon sa balikat ng palaka:
1 - presternum; 2 - clavicle; 3 - coracoid (buto ng uwak); 4 - sternum; 5 - scapula at suprascapular cartilage.
na may malalaking suprascapular cartilages, crow bones at collarbones, pati na rin ang isang unpaired bone - ang sternum (Fig. 146). Ang pelvic girdle ay binubuo ng tatlong magkapares na buto, na pinagsama dahil sa mabibigat na karga: iliac, pubic at ischial. Sa tulong ng ilium, ang pelvic girdle ay nakakabit sa mga transverse na proseso ng sacral vertebrae.
Dahil sa mas kumplikadong paggalaw, ang muscular system sa amphibians ay mas kumplikado kaysa sa isda. Ang makapangyarihan at kumplikadong organisadong mga kalamnan ng unahan at hulihan ay bubuo. Ang metamerism ng kalamnan (segmented na istraktura), katangian ng isda, ay nabalisa sa mga amphibian, ang istraktura ng muscular system ay nagiging mas naiiba.
Sistema ng pagtunaw
nailalarawan sa pamamagitan ng pagkuha ng ilang mga tampok na nauugnay sa isang terrestrial na paraan ng pamumuhay. Hindi tulad ng mga isda, ang mga amphibian ay nagkakaroon ng mga glandula ng salivary, na ang mga duct ay bumubukas sa oropharyngeal cavity. Bilang karagdagan, lumilitaw ang isang dila, na may sariling mga kalamnan at nakikibahagi sa pagkuha at pagpapanatili ng pagkain. 
kanin. 147. Ulo ng palaka:
1 - laryngeal fissure; 2 - wika; 3, 4 - panloob at panlabas na butas ng ilong, ayon sa pagkakabanggit; 5 - mata; 6 - tympanic membrane.
Ang dila ay matatagpuan sa ilalim ng oropharyngeal cavity at maaaring magkaroon ng ibang hugis. Sa palaka, ang likod ng dila ay maaaring ihagis pasulong at nagsisilbing kasangkapan sa paghuli ng maliliit na hayop (Larawan 147). Ang maliliit na ngipin sa itaas na panga, na may hugis na korteng kono at bahagyang hubog sa likod na tuktok, ay nakakatulong din sa pagpapanatili ng pagkain; Ang pagtulak sa bolus ng pagkain ay pinadali ng mga eyeballs, na nahihiwalay mula sa oropharyngeal na lukab lamang ng isang manipis na mucous membrane at maaaring iguguhit sa oropharynx sa tulong ng mga espesyal na kalamnan.
Fig.148. Diagram ng istraktura ng baga ng palaka.
ang paghahanap mula sa oropharynx sa pamamagitan ng isang maikling esophagus ay pumapasok sa tiyan. Ang bituka ay medyo mas mahaba kaysa sa isda, at binubuo ng tatlong seksyon: anterior, middle, at posterior. Ang tumbong ay bumubukas sa cloaca. Ang malaking atay ay naglalabas ng apdo, na naipon sa gallbladder at pumapasok sa mga duct sa nauunang bahagi ng maliit na bituka (ang tinatawag na duodenum), kung saan dumadaloy din ang mga pancreatic duct.
Sistema ng paghinga . Ang amphibian larvae ay humihinga sa tulong ng mga branched na panlabas na hasang, na nawawala pagkatapos ng metamorphosis sa karamihan ng mga species. Sa mga amphibian na nasa hustong gulang, ang palitan ng gas ay nangyayari sa pamamagitan ng balat at baga. Ang halaga ng paghinga ng balat ay napakataas, halimbawa, sa isang berdeng palaka, 51% ng oxygen ang pumapasok sa balat at 86% ng carbon dioxide ay inilabas.
Ang mga baga ng amphibian ay mga guwang na bag na may mas marami o hindi gaanong binibigkas na cellular na istraktura (Larawan 148). Ang ibabaw ng baga ay napakaliit, at ang ratio nito sa ibabaw ng balat ay 2:3. Ang respiratory tract ay hindi rin maayos na binuo. Sa isang palaka, sila ay kinakatawan ng isang maikling tracheal-laryngeal chamber, at sa mga kinatawan ng caudates, sila ay kinakatawan ng isang mahabang tubo - ang trachea.
kasi rib cage wala, ang mekanismo ng paghinga ay isang primitive forced type. Ang paglanghap ay isinasagawa sa pamamagitan ng mga butas ng ilong, habang ibinababa ang ilalim ng bibig, pagkatapos ay ang mga butas ng ilong ay sarado na may mga balbula, ang ilalim ng bibig ay tumataas muli, at ang hangin ay itinulak sa mga baga. Ang pagbuga ay nangyayari sa tulong ng mga kalamnan ng tiyan. Bilang karagdagan sa mga baga, ang karagdagang gas exchange ay isinasagawa sa oral cavity, ang pader na kung saan ay natagos ng isang network ng mga capillary.
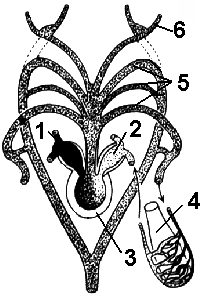
kanin. 149. Scheme ng istraktura ng puso at arterial arches:
1 - kanang atrium; 2 - kaliwang atrium; 3 - ventricle; 4 - baga; 5 - arterial arches; 6 - carotid arteries.
ov.
Daluyan ng dugo sa katawan nahahati sa dalawang bilog ng sirkulasyon ng dugo: malaki at maliit (pulmonary). Ang puso ay may tatlong silid, na nabuo ng dalawang atria at isang ventricle. Ang arterial blood ay pumapasok sa kaliwang atrium mula sa mga baga sa pamamagitan ng pulmonary veins, at ang halo-halong dugo ay pumapasok sa kanang atrium, dahil ang vena cava ay mula sa lamang loob pumapasok ang venous blood, at ang mga ugat ng balat ay nagdadala ng arterial blood (Fig. 149).
Sa ventricle, ang dugo ay bahagyang naghahalo, dahil sa pagkakaroon ng mga espesyal na mekanismo ng paghahati (iba't ibang mga outgrowth at isang spiral valve ng arterial cone).
Mahusay na bilog ng sirkulasyon ng dugo. Mula sa ventricle, ang dugo ay pumapasok sa arterial cone, na nagsasanga pa sa tatlong pares ng arterial vessel. Kapag nagkontrata ang ventricle, ang venous blood ay unang itinutulak palabas, na pumupuno sa unang dalawang pares ng mga arterya. Ang dugo na may pinakamataas na nilalaman ng oxygen ay pumapasok sa ikatlong pares ng mga arterya, kung saan umaalis ang mga carotid arteries, na nagbibigay ng dugo sa utak.
Ang mga arko ng aorta, na inilarawan ang isang kalahating bilog, ay pinagsama at bumubuo ng isang karaniwang puno ng dorsal aorta, na nagbibigay ng mga panloob na organo na may halo-halong dugo. Pagkatapos, ang venous blood (mula sa mga panloob na organo sa pamamagitan ng vena cava) at arterial (sa pamamagitan ng mga ugat ng balat) ay pumasok sa kanang atrium.
Ang pulmonary arteries ay nagdadala ng oxygen-poor blood sa baga, kung saan nangyayari ang palitan ng gas, pagkatapos ay sa pamamagitan ng pulmonary veins, ang arterial blood ay pumapasok sa kaliwang atrium - ito ang pulmonary circulation. Ang mga malalaking sanga ay umaalis sa bawat pulmonary artery - mga arterya ng balat na nagdadala ng dugo sa balat, kung saan ito ay na-oxidized, at pagkatapos ay pumapasok sa kanang atrium.
Ang mga erythrocytes sa amphibian ay malaki, biconvex, may nucleus. Ang metabolismo ay mas mataas kaysa sa isda, ngunit hindi sapat na mataas upang mapanatili ang isang pare-parehong temperatura ng katawan, kaya ang mga amphibian ay inuri bilang mga poikilothermic na hayop.
H
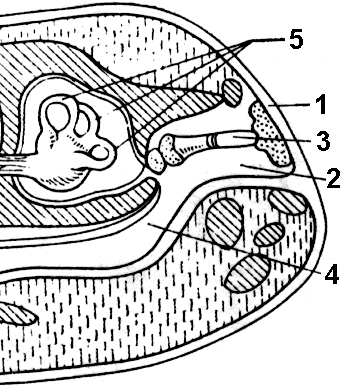
Figure 150. Cross section sa ulo ng palaka sa bahagi ng tainga:
1 - eardrum; 2 - lukab ng gitnang tainga; 3 - stirrup; 4 - Eustachian tube; 5 - kalahating bilog na mga kanal.
Sistema ng nerbiyos at mga organo ng pandama. Ang utak ng mga amphibian, tulad ng sa lahat ng vertebrates, ay binubuo ng limang seksyon: ang forebrain, diencephalon, midbrain, cerebellum at medulla oblongata. Kung ikukumpara sa isda, ang relatibong laki ng forebrain ay tumaas sa mga amphibian at nagkaroon ng kumpletong paghihiwalay ng mga hemisphere nito. Bilang karagdagan, ang bilang ng mga selula ng nerbiyos (grey matter) ay tumaas, na, gayunpaman, ay nakapaloob sa malalim na mga layer ng forebrain at wala sa ibabaw nito. Ang midbrain ay medyo maliit, at ang cerebellum ay kulang sa pag-unlad dahil sa mababang mobility at monotonous na paggalaw. Ang medulla oblongata ay dumadaan sa spinal cord.
Organ ng pandinig. Bilang karagdagan sa panloob na tainga, na matatagpuan din sa isda, ang mga amphibian ay may binuo na gitnang tainga, sa lukab kung saan mayroong isang stirrup - isang pandinig na buto, na unang lumitaw sa mga vertebrates.
Upang magpadala ng mga tunog na vibrations, ang stirrup sa isang dulo ay nakasalalay sa tympanic membrane, na naghihiwalay sa gitnang tainga na lukab mula sa panlabas na kapaligiran (Larawan 150), at sa kabilang dulo laban sa hugis-itlog na bintana, na isang manipis na seksyon ng septum sa pagitan ng gitna at panloob na tainga. Ang gitnang tainga na lukab ay konektado sa oral cavity sa pamamagitan ng isang makitid na kanal - ang Eustachian tube, ito ay kinakailangan upang ang panlabas at panloob na mga presyon sa tympanic membrane ay pareho, na nagpoprotekta sa lamad mula sa pagkalagot.
Ang organ ng paningin ay kinakailangan para sa mga amphibian upang suriin ang mga bagay, lalo na sa lupa. Ang mga adaptasyon na kinakailangan para dito ay ipinahayag sa matambok na hugis ng kornea at sa lens, na nasa anyo ng isang biconvex lens (sa kaibahan sa isda, na may spherical lens). Bilang karagdagan, upang maprotektahan ang mga mata mula sa pagbara at pagkatuyo, ang mga amphibian ay may mga movable eyelids, nictitating membrane, at mga glandula na nagmo-moisturize sa cornea. Ang tirahan ay nangyayari, tulad ng sa isda, dahil lamang sa paggalaw ng lens.
Ang olfactory organ ng amphibians ay matatagpuan sa mga ipinares na olfactory capsule, na nakikipag-usap sa panlabas na kapaligiran sa pamamagitan ng mga panlabas na butas ng ilong, at sa oropharyngeal na lukab - sa pamamagitan ng choanae, sa gayon, ang inilarawan na sistema ay nagsisilbi hindi lamang para sa pang-unawa ng mga amoy, kundi pati na rin para sa paghinga. Ang lateral line ay katangian ng lahat ng amphibian larvae at matatagpuan sa balat, ngunit hindi sa isang deepened channel, tulad ng sa isda, ngunit mababaw. Sa mga pormang pang-adulto, ang lateral line ay pinapanatili lamang sa mga aquatic tailed amphibian at ilang anuran, na aquatic din.
excretory system , tulad ng sa isda, ito ay kinakatawan ng dalawang trunk kidney, ang pag-andar nito ay upang alisin ang labis na tubig. Ang pangunahing produkto ng excretion ay urea. Ang mga ureter ay nagdadala ng ihi sa cloaca at pagkatapos ay sa pantog. Pagkatapos punan ito, ang ihi ay ilalabas muli sa cloaca, at pagkatapos ay lumabas. Sa mga lalaki, ang ureter ay gumaganap ng isa pang function - ang function ng vas deferens.
Ang mga amphibian ay mga hyperosmotic na hayop na may paggalang sa sariwang tubig. Bilang isang resulta, ang tubig ay patuloy na pumapasok sa katawan sa pamamagitan ng balat, na walang mga mekanismo upang maiwasan ito, tulad ng sa iba pang mga terrestrial vertebrates. Ang tubig sa dagat ay hyperosmotic na may kaugnayan sa osmotic pressure sa mga tisyu ng amphibian, ang tubig ay umaalis sa katawan sa pamamagitan ng balat. Ito ang pinakamahalagang dahilan kung bakit hindi mabubuhay ang mga amphibian sa tubig ng dagat at mamatay dito dahil sa dehydration.
Pagpaparami at pag-unlad . Ang mga pagkakaiba sa kasarian sa pagitan ng mga babae at lalaki ng mga amphibian ay kadalasang hindi maganda ang pagpapahayag. Sa karamihan ng mga anuran, ang mga lalaki ay medyo mas maliit kaysa sa mga babae. Ang mga male newts ay may maliwanag na kulay ng isinangkot. May mga pagbabago din morphological character, halimbawa, sa male crested newt, sa panahon ng pag-aanak, lumalaki ang isang leathery rim sa likod at buntot, at sa mga lalaki ng tailless amphibian, isang nuptial callus ang nabubuo sa loob ng front legs.
Ang pagpapabunga ay maaaring maging panloob at panlabas.
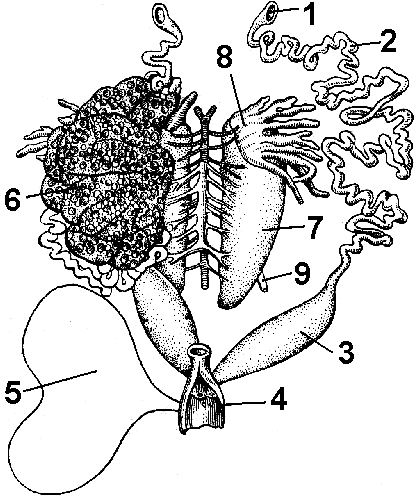
kanin. 151. Urogenital system ng babaeng palaka:
1 - funnel ng oviduct; 2 - oviduct (Mullerian canal); 3 - matris; 4 - cloaca; 5 - pantog; 6 - obaryo; 7 - bato; 8 - taba ng katawan; 9 - yuriter.
pangit. Ang pag-unlad ng mga amphibian ay nangyayari sa metamorphosis, iyon ay, ang larvae ay lumilitaw mula sa mga itlog na karaniwang nabubuo sa tubig. Pinapakain nila ang pagkain na iba sa diyeta ng mga matatanda at naiiba sa mga matatanda sa ilang mga tampok na istruktura na tulad ng isda.
Ang mga genital organ ng lalaking palaka ay kinakatawan ng magkapares na testes. Ang seminiferous tubules ay walang laman sa ureter (Wolf's canal). Ang pagpapabunga sa palaka ay panlabas, ayon sa pagkakabanggit, ang lalaki ay walang copulatory organs.
Ang reproductive system ng babae ay kinakatawan ng mga magkapares na ovaries at oviducts (Müllerian canals), na mukhang napakagulong mga tubo, na dumadaloy sa cloaca sa isang dulo, at nagdadala ng mga funnel sa kabilang dulo, kung saan nahuhulog ang mga itlog (Fig. 151).
Ang pula ng itlog sa itlog ay hindi pantay na ipinamamahagi, ito ay puro sa mas mababang vegetative plus ng isang mas magaan na kulay. Ang tapat (poste ng hayop) ay naglalaman ng mga itlog
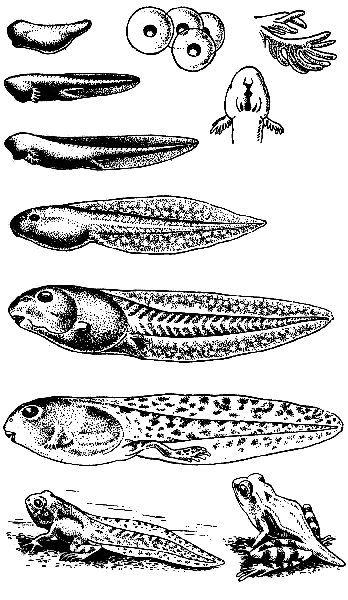
Fig.152. Pag-unlad ng palaka.
ito ay isang pigment na nagbibigay ito ng isang mas madilim na kulay, na nagsisilbi para sa mas mahusay na pag-init.
Ang itlog ay nakakaranas ng kumpleto at hindi pantay na pagdurog. 8-10 araw pagkatapos ng pagpapabunga, ang embryo ng palaka ay pumutok sa mga kabibi ng itlog at sa anyo ng isang larva - isang tadpole - ay lumalabas.
Sa una, ang tadpole ay may istraktura na tulad ng isda: wala itong ipinares na mga paa, ang tanging organ ng paggalaw ay ang buntot na may mahusay na nabuo na lamad. Ang mga organ ng paghinga nito ay 2-3 pares ng mga panlabas na hasang (Larawan 152), ang lateral line ay mahusay na binuo, mayroong isang chord, apat na arterial arches. Sa yugtong ito ng pag-unlad, ang tadpole ay may dalawang silid na puso at isang bilog lamang ng sirkulasyon ng dugo, tulad ng sa isda.
Sa karagdagang pag-unlad, ang mga panlabas na hasang atrophy at hasang slits na may petals form, na kung saan din pagkatapos ay nawawala habang ang mga baga ay nabuo. Kaayon nito, ang iba pang mga proseso ay nangyayari din: ang lateral line ay nawawala, ang buntot ay atrophies, ipinares na mga limbs ay nabuo, ang notochord ay nawawala, at ang mga bituka ay umiikli. Ang tadpole ay dumadaan mula sa pagkain ng halaman hanggang sa pagkain ng hayop, na katangian ng mga matatanda, ay nagiging palaka.
Order Tailed amphibians (Caudata). Ang pinaka sinaunang at medyo maliit na grupo ng mga amphibian - mga 340 species. Ang mga ito ay kadalasang ipinamamahagi sa temperate zone ng kanluran at silangang hemisphere. Ang katawan ay pinahaba, bilugan, na may mahabang buntot na nagpapatuloy sa buong buhay. Magkapareho ang haba ng unahan at hulihan, kaya gumagapang o naglalakad ang mga ito. Ang bisig at ibabang binti ay may tipikal na istraktura at binubuo ng dalawang buto, ang ulna at radius, ang tibia at tibia. Mga tampok panloob na istraktura ay ang pinaka-primitive para sa klase sa kabuuan. Sa pinaka-primitive na mga anyo, ang isang panimulang chord ay pinapanatili sa buong buhay. May mga panimulang itaas na tadyang.
Ang ilang mga kinatawan, tulad ng Proteus, ay nagpapanatili ng mga panlabas na hasang sa buong buhay nila. Ang papel ng paghinga ng balat ay mahusay. Ang lukab ng gitnang tainga at ang tympanic membrane ay wala; sa ilang mga species, ang lateral line ay napanatili habang buhay. Ang pagpapabunga sa karamihan ng mga species ay panloob, kinukuha ng babae ang mga mucous sac na may spermatozoa (spermatophores) kasama ang cloaca.
Ang ilan ay naobserbahan ang proseso ng pagpaparami sa yugto ng larval - neoteny. Ang hindi pangkaraniwang bagay na ito ay pinakamahusay na kilala para sa axolotl, isang North American amphibian na may kakayahang magparami ngunit talagang ang larval stage ng ambystoma.
Mag-order ng mga amphibian na walang buntot (Anura). Walang buntot - ang pinaka-mataas na organisado at maraming grupo ng mga amphibian, na kasalukuyang may bilang na 2900 species. Sa lahat ng iba't ibang uri ng hayop, panlabas na istraktura medyo monotonous, na nauugnay sa paggalaw sa pamamagitan ng paglukso. Ang hugis ng katawan ay pinaikli, hugis-wedge. Ang mga hind limbs ay pinahaba, isang bagong seksyon ang lilitaw sa kanila, ang "pangalawang ibabang binti", na nabuo sa pamamagitan ng pagpapahaba ng dalawang buto ng tarsus. Dapat tandaan na ang mga nakapares na buto sa ibabang binti at bisig ay pinagsama para sa mas mahusay na suporta kapag gumagalaw sa lupa.
Ang detatsment ay nahahati sa 19 na pamilya, ang pinakasikat sa mga ito ay: ang Round-tongued family (toads), ang Pipaceae family (spur frog, Surinamese pipa), ang Spadefoot family (common spadefoot), ang Real toads family (gray at green toads), ang Frog family (common tree frog ), isang pamilya ng mga tunay na palaka (lawa at pond frog).
Mga pangunahing termino at konsepto
1. Spiral valve. 2. Mga bato ng puno ng kahoy. 3. Ureothelium. 4. Lobo channel. 5. channel ni Müller. 6. Gitnang tainga. 7. Eustachian tube. 8 Neoteny. 9. Ripidistia. 10. Ichthyostegids. 11. Stegocephaly.
Mahahalagang Tanong sa Pagsusuri
Anong mga order ang nahahati sa modernong amphibian?
Anong mga tampok ang nakuha ng mga amphibian kaugnay ng landfall?
Anong mga departamento ang nakikilala sa anterior at hind limbs?
Anong mga departamento ang nakikilala sa gulugod?
Ano ang balikat at pelvic girdle ng limbs?
Aling mga pangunahing glandula ng pagtunaw ang nagbubukas sa duodenum?
Ano ang pangalan ng huling seksyon ng bituka, kung saan nagbubukas ang malaking bituka, excretory at reproductive system?
Ano ang mga respiratory organ ng larvae at adult amphibian?
Gaano karaming mga bilog ng sirkulasyon ng dugo at kung gaano karaming mga silid ang nasa puso ng mga adult amphibian?
Gaano karaming mga bilog ng sirkulasyon ng dugo at kung gaano karaming mga silid ang nasa puso ng isang tadpole?
Anong dugo ang pumapasok sa kanan at kaliwang atrium?
Anong mga organo ang bumubuo excretory system amphibian?
Ano ang mga bato ng mga adult amphibian?
Sa anong anyo ang mga produkto ng metabolismo ng nitrogen ay pinalabas sa mga amphibian ng may sapat na gulang?
Sa anong anyo ang mga produkto ng nitrogen metabolism ay pinalabas sa amphibian larvae?
Paano naiiba ang pandinig at paningin ng mga amphibian sa mga isda?
Anong mga tampok ang nagpapahintulot sa mga isda na makabisado ang terrestrial na tirahan at maging mga amphibian?
Phylogeny ng mga amphibian.
Ang lahat ng mga vertebrates ay may dalubhasang mga organ sa paghinga (maliban sa ilang amphibian, na nawala ang mga ito sa pangalawang pagkakataon). Ang mga organo na ito, bilang panuntunan, ay bubuo mula sa mga paglaki ng tubo ng bituka (sa itaas, ang phylogenetic na relasyon sa pagitan ng mga pag-andar ng paghinga at nutrisyon ay ipinahiwatig).
Ang pangkalahatang prinsipyo ng paggana ng mga sistema ng paghinga ay upang lumikha dynamic na contact ng dugo na may gaseous medium, sa kumbinasyon ng daloy ng dugo sa mga sisidlan ng mga organ ng paghinga (ang kanilang perfusion) na may pumping ng tubig o hangin sa pamamagitan ng mga organ na ito (ang kanilang bentilasyon). Ang isang pagbubukod ay ang paghinga ng balat ng mga amphibian, na hindi nangangailangan ng isang espesyal na kagamitan sa bentilasyon.
Paghinga ng tubig.Ang paghinga ng tubig sa tulong ng mga hasang ay likas sa karamihan ng isda. Ang mga hayop na ito ay kumukuha ng tubig sa oral cavity, pagkatapos nito ay itinutulak palabas ang tubig sa pamamagitan ng mga gill slits. Ginagamit sa pagkuha ng oxygen prinsipyo ng counterflow(Larawan 10.4). Ang dugo sa mga capillary ng hasang ay dumadaloy patungo sa tubig na naghuhugas ng mga gill sheet, na pinadali ng paggalaw ng mga isda sa kalawakan. Ang mataas na kahusayan ng paghinga ng hasang ay idinisenyo upang mabayaran ang malaking paggasta ng enerhiya sa gawain ng mga kalamnan na nagpapahangin sa mga hasang gamit ang tubig. Ang kalamnan na ito ay kinokontrol ng isang hiwalay sentro ng paghinga
|
kanin. 10.4. Gill arches (apat) sa isda. (A), isang fragment ng dalawang gill arch (B), isang seksyon ng gill filament na may tatlong gill filament sa bawat gilid (C), at isang seksyon ng bony fish hasang kung saan ang dugo at tubig ay dumadaloy sa magkasalungat na direksyon (D) 1 - takip ng gill, 2 - constrictor na kalamnan, 3 - gill filament, 4 - gill cavity, 5 - mga daluyan ng dugo, 6 - oral cavity, 7 - gill skeleton, 8 - plates, 9 - cross section sa pamamagitan ng gill filament, 10 - septum, 11 - septal canal, 12 - gilid ng plate. |
Vmedulla oblongata, papalapit sa samahan nito sa respiratory center ng mga mammal. Pagtuklas ng ritmikong aktibidad ng mga istruktura sa nakahiwalay na brainstem ng isang goldpis ( Carassius auratus auratus) ay isang mahalagang milestone sa kaalaman automatism ng respiratory center.
Dahil sa mataas na solubility ng CO 2 sa tubig, madali itong inilabas, kaya ang gawain ng respiratory apparatus ng isda ay pangunahing nagsisilbi upang magbigay ng oxygen sa katawan. Ang mga senyales tungkol sa nilalaman ng O2 sa dugo, na nagmumula sa mga chemoreceptor ng mga daluyan ng hasang, ay hindi lamang kinokontrol ang aktibidad ng respiratory center ng isda, ngunit hinihimok din itong pumili ng mga zone ng reservoir na may physiologically optimal na oxygenation. Ang pag-uugaling tugon na ito ng aktibong pagpili ng isang ginustong kapaligiran sa paghinga ay tinatawag kagustuhan sa gas. Ang ilang mga isda (eels) ay sumisipsip ng oxygen na natunaw sa tubig pangunahin hindi sa pamamagitan ng mga hasang, ngunit gamit ang pagsasabog nito sa pamamagitan ng balat.
Paglipat mula sa tubig tungo sa paghinga ng hangin. Maraming mga isda, na paminsan-minsang umaakyat sa ibabaw ng mga anyong tubig o kahit na lumalapag at lumulunok ng hangin, ay nagdaragdag ng tubig sa paghinga ng hangin, kung saan ginagamit nila ang mga pader na may malaking vascularized ng oral cavity, pharynx, tiyan, bituka, o isang espesyal na derivative ng ang tubo ng bituka - paglangoy pantog.
Lungfish. huminga na ng hangin sa tulong ng isang bagong dalubhasang organ - baga na isang nakatiklop na sako (single o paired) na nagmumula sa ventral protrusion ng pharynx. Ilang species ng bipedal fish ( Lepidosireniformes) nakatira sa tropikal na tubig, mahina ang aerated at acidified din. Hindi nila magagawa nang walang paghinga ng hangin.
|
|
|
kanin. 10.5 Ang prinsipyo ng paggana ng mga sistema ng paghinga ng mga vertebrates 1 - dugo, 2 - hadlang, 3 - kapaligiran, 4 - mga organ sa paghinga, 5 - pagsasabog; Pi, Re - bahagyang presyon ng O 2 sa inhaled at exhaled hangin (tubig); Pa, Pv- pareho sa arterial at venous blood. |
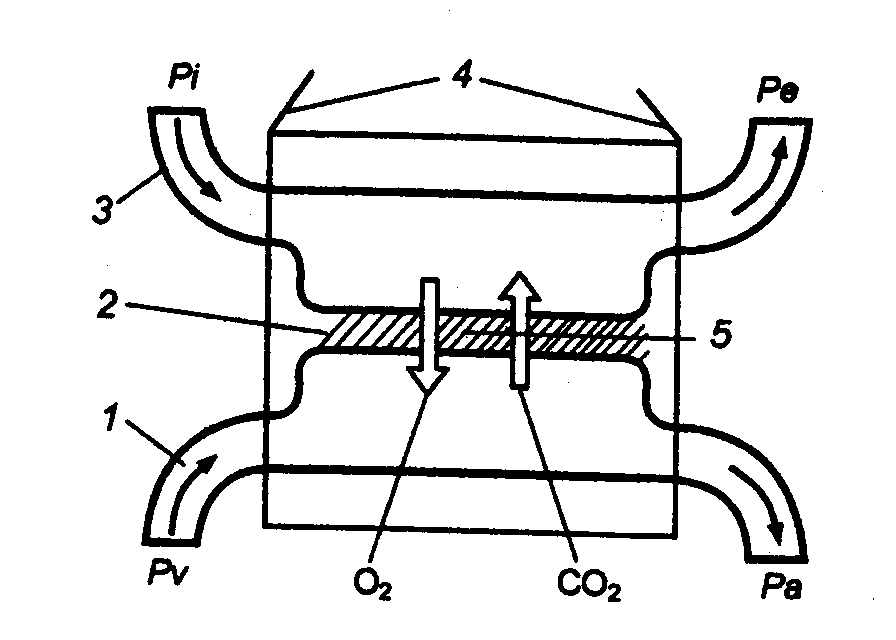
Ang ganitong pagbabago sa mga kondisyon, malinaw naman, ay nag-ambag sa mass transition ng aquatic vertebrates (sila ay lobe-finned fish - ang mga ninuno ng mga amphibian) sa hangin na paghinga sa panahon ng Devonian (mga 400 milyong taon na ang nakalilipas). Dapat itong isaalang-alang na sa oras na iyon ang bahagyang presyon ng 02 sa kapaligiran ay malapit sa modernong isa. At kung ang 1 litro ng tubig na puspos ng hangin sa temperatura na 0 ° C ay naglalaman lamang ng 0.014 g 02 (sa 30 ° C ang halagang ito ay nahahati), kung gayon sa 1 litro ng hangin ang nilalaman nito ay 0.28 g, ibig sabihin, sa 20 beses na higit pa. . Bilang karagdagan, ang density ng hangin ay higit sa 700 beses na mas mababa kaysa sa tubig, kaya ang bentilasyon nito sa pamamagitan ng mga organ ng paghinga ay nangangailangan ng katumbas na mas kaunting pagkonsumo ng enerhiya. Karamihan sa mga amphibian (hindi bababa sa kanilang mga pang-adultong anyo) ay lumipat sa paghinga ng hangin, bagaman ang mga baga ng mga amphibian ay hindi pa rin nabuo: ang ibabaw ng gas exchange ay maliit, ang venous at arterial na daloy ng dugo sa sistema ng sirkulasyon ay hindi ganap na natunaw. Ang ilan sa mga hinihigop na O 2 at karamihan sa mga ibinubuga na CO 2 ay hindi kumakalat sa pamamagitan ng mga baga, ngunit sa pamamagitan ng moistened na balat, ang vascularization kung saan nagbabago alinsunod sa intensity ng skin gas exchange (Fig. 10.5).
Mga paggalaw ng paghinga amphibian(tulad ng mga reptilya) ay katangian - sa ito ay naiiba sila sa parehong isda at mammal - mahabang paghinto, apnea, sa panahon kung saan tanging ang buccal-pharyngeal na lukab ay maaliwalas na ang glottis ay sarado, na naghihiwalay sa mga baga mula sa atmospera. Paminsan-minsan lamang mayroong mga coordinated contraction ng mga kalamnan ng bibig at larynx, na pinipilit ang hangin sa mga baga at sinisipsip ito pabalik. Ang mga paggalaw na ito ay kinokontrol ng mga nerve center ng brainstem alinsunod sa mga impulses ng mga stretch receptor ng baga, pati na rin ang mga arterial chemoreceptors, na nasasabik sa kakulangan ng oxygen sa dugo. Bilang karagdagan, maraming amphibian ang nagpapakita ng regulasyon sa pag-uugali ng paghinga, isang kagustuhan sa gas: depende sa mga kondisyon, pinipili nila ang alinman sa isang aquatic o isang kapaligiran sa hangin.
Terrestrial mga reptilya may kaugnayan sa pagbuo ng takip ng keratinizing, ang paghinga ng tubig at balat ay nagiging imposible (maliban sa mga butiki). Ang yugtong ito sa ebolusyon ng paghinga ay nagmamarka ng pangwakas na paglipat mula sa palitan ng gas sa isang mahalumigmig na kapaligiran hanggang sa "tuyo" na bentilasyon, ang mekanismo na kung saan ay halos kapareho ng sa mga mammal. Ang ibabaw ng gas exchange ng mga baga ay makabuluhang nadagdagan dahil sa pagbuo ng mga maliliit na selula, ang dugo ng isang nakahiwalay na maliit na bilog ay dumadaloy sa isang siksik na network ng kanilang mga sisidlan. Kasabay nito, ang mga reptilya ay nagpapanatili pa rin ng paulit-ulit na likas na bentilasyon ng mga baga, na puno ng hangin, gayunpaman, sa pamamagitan ng hindi pagpilit, ngunit sa pamamagitan ng pagsuso nito.
Metabolismo ectothermic (poikilothermic) vertebrates ay medyo mababa pa rin, na siyang dahilan ng medyo maliit na "kapasidad" ng kanilang respiratory apparatus. Ayon sa ilang mga mananaliksik, ito ang dahilan ng kawalan ng kakayahan ng mga wala na ngayong malalaking reptilya na umangkop sa mga dramatikong pagbabago sa tirahan na naganap sa pagtatapos ng Mesozoic.
Makabuluhang mas matindi at matatag na mga proseso ng metabolic sa endotherms (homeothermic na hayop) - mga mammal at ibon. Mayroong karagdagang pagtaas sa ibabaw ng gas exchange at suplay ng dugo sa mga baga. Hindi tulad ng mga amphibian at reptilya, isang bagong nabuo maindayog, walang paghinto, uri ng bentilasyon ng baga. Makabuluhang nagbabago ang mekanismo ng regulasyon ng paghinga dahil sa ang katunayan na ang kabuuan CO2 sa mga hayop na ito, halos tinatanggal lamang ito ng mga baga, at ang produksyon nito ay nagiging napakatindi dahil sa pagtindi ng mga proseso ng oxidative. May kahirapan sa paglaban sa akumulasyon ng carbonic acid sa mga tisyu at dugo. Samakatuwid, sa mga hayop na eksklusibong humihinga gamit ang mga baga, kahit na ang arterial blood pressure ay tumataas, ang Pco 2 ay tumataas nang malaki sa parehong oras (Fig. 10.6), at ang kapangyarihan ng mga buffer system ay tumataas din.
|
|
|
kanin. 10.6 Pag-igting ng respiratory gas sa arterial blood (PaO 2 P a co 2 ) sa iba't ibang klase vertebrates Ang paghinga ng hasang ng isda ay nagbibigay sa kanila ng oxygen; R co 2 mababa ang dugo nila, kasi co 2 madaling kumalat sa pamamagitan ng integument ng katawan sa nakapalibot na tubig. Ang pag-uugali ng paghinga at paghinga ng hasang sa isda ay kinokontrol ng oxygen. Para sa mga isda na iyon ilang kundisyon gumamit ng hangin na paghinga (pati na rin sa mga reptilya at mammal), hindi lamang ang paggamit ng O 2, kundi pati na rin ang pagbabalik co 2 sa hangin ay nangyayari lamang sa pamamagitan ng mga baga. Samakatuwid R co 2 ang dugo sa mga hayop na humihinga ng hangin ay mas mataas kaysa sa mga hayop na humihinga ng tubig, at ang parameter na ito ay nagiging pangunahing regulator ng paghinga. Gayunpaman, si R o 2 ang dugo sa tubig- at hangin-breathers ay bahagyang naiiba. |

Dahil sa mga pangyayaring ito, ang papel ng pangunahing regulator ng paghinga ay pumasa mula sa O 2 hanggang CO 2 at ang mga H + ions na nauugnay sa transportasyon nito, at ang mga sentral (medullary) ay nakakakuha ng nangungunang papel sa pagpapasigla ng ritmikong aktibidad ng respiratory center. sa halip na arterial chemoreceptors. mga chemosensitive na lugar. Ang lahat ng mga tampok na ito ay saklaw sa mga nauugnay na seksyon ng kabanata. Kasabay nito, ang paghinga ng mga ibon ay ibang-iba sa paghinga ng mga mammal na dapat nating pag-isipan ito nang hiwalay.
Hininga ng mga ibon.Ang mga baga ng mga ibon, na may spongy na istraktura, ay naiiba, una, sa ang palitan ng gas sa pagitan ng hangin at dugo ay nangyayari sa tinatawag na air capillaries, "cylindrical alveoli" (sa pamamagitan ng pagkakatulad sa saccular alveoli ng mga mammal) - at, pangalawa , sa sistema ng daloy ng bronchial. Ang sistemang ito ay dinisenyo sa paraang ang mga baga, sa tulong ng ilan Lagayan ng hangin ay maaliwalas ng sariwang hangin kapwa kapag humihinga at humihinga (tingnan ang Fig. 10.7). Ang ganitong uri ng paghinga ay nagbibigay sa pulmonary gas exchange ng ibon ng mataas na kahusayan na kinakailangan upang masakop ang masinsinang paggasta ng enerhiya, lalo na sa paglipad. Ang mga paggalaw ng paghinga ng pader ng dibdib ay nilikha pangunahin sa pamamagitan ng mga contraction ng mga kalamnan ng pectoral at tiyan. Sa paglipad, ang bentilasyon ng mga baga (na, bilang karagdagan sa palitan ng gas, ay kasangkot sa pagpapalabas ng labis na init) ay nakakatulong sa paggalaw ng mga pakpak.
Ang sentro ng paghinga ng ibon ay matatagpuan sa medulla oblongata. Ang aktibidad nito, tulad ng sa mga mammal, ay kinokontrol ng mga impulses mula sa medullary chemoreceptors na sensitibo sa CO 2 at H + ions, pati na rin ang mga arterial chemoreceptor na tumutugon din sa kakulangan ng O 2, na mahalaga para sa regulasyon ng paghinga habang lumilipad. sa malalaking taas. Sa mga daanan ng hangin ng ibon mayroong mga mechanoreceptor, ang pagpapasigla nito ay kasangkot sa regulasyon ng dalas at lalim ng paghinga. Mayroon ding mga espesyal na chemoreceptor, ang impulsasyon na tumataas sa pagbawas sa konsentrasyon ng intrapulmonary CO2 . Ang pagpapasigla ng mga receptor na ito ay pumipigil sa aktibidad ng respiratory center at sa gayon ay binabawasan ang bentilasyon ng baga, na pumipigil sa labis na paglabas ng CO 2 mula sa katawan, na maaaring mangyari sa ilalim ng mga kondisyon ng paglipad.