Семязачатки - это небольшие образования, располагающиеся в завязи пестика и развивающиеся в семя. Семязачаток формируется
из меристематического бугорка, возникающего на плодолистике. Место прикрепления семязачатка к плодолистику называется пла- центой. Число семязачатков может колебаться от одного (пшеница, вишня, подсолнечник) до нескольких тысяч (мак) и миллионов (у орхидных).
Развившийся семязачаток имеет семяножку, посредством которой он прикрепляется к плаценте; центральную часть - нуцеллус (гомолог мегаспорангия); 1 или 2 интегумента (покрова) семязачатка, которые на верхушке не срастаются и образуют микропиле, или пыльцевход; противоположную микропиле базальную часть семязачатка - халазу, где нуцеллус и интегументы сливаются. В семязачатке происходят следующие процессы: мегаспорогенез - формирование мегаспор; мегагаметогенез - формирование женского гаметофита и оплодотворение.
Микроспорогенез и микрогаметогенез
Микроспорангиями являются гнезда пыльников. Процесс образования микроспор в микроспорангиях называется микроспорогене- зом. Клетки спорогенной ткани, в гнездах пыльников, в результате митотических делений формируют материнские клетки микроспор (микроспороциты). Из каждого диплоидного микроспороцита в результате мейоза образуются 4 гаплоидные микроспоры. Сформировавшаяся микроспора имеет оболочку и единственное гаплоидное ядро. Такая микроспора дает начало пыльцевому зерну. Процесс образования из микроспоры мужского гаметофита - пыльцы - называется микрогаметогенезом.
Микрогаметогенез. При делении митозом из микроспоры формируется пыльца, состоящая из 2 новых клеток и покрытая оболочкой - спородермой. Маленькая клетка - генеративная (спермагенная), а более крупная - вегетативная (клетка пыльцевой трубки). Таким образом, пыльца, или мужской гаметофит, формируется из микроспоры внутри пыльника.
Спородерма пыльцы имеет 2 слоя - экзину и интину. Наружная слоистая экзина пропитана очень стойкими углеводами спорополленинами и имеет на поверхности различные выросты. Внутренняя оболочка - интина. Она менее стойка, чем экзина, состоит из целлюлозы и пектина и сохраняет эластичность (рис. 4.8). Особенности строения спородермы и пыльцы являются видоспецифичными. После созревания пыльцы пыльники лопаются, и пыльца рассеивается.
Рис. 4.8. Микроспора и ее прорастание: А - микроспора; Б - пыльцевое зерно; В - формирование пыльцевой трубки; Г - часть пыльцевой трубки; 1 - экзина; 2 - интина; 3 - вегетативная клетка; 4 - репродуктивная клетка; 5 - спермии
Жизнеспособность пыльцы может сохраняться от нескольких часов (у ржи - 12 ч) до нескольких лет (у яблони 4,5 года).
Мегаспорогенез и мегагаметогенез
Центральной частью семязачатка является нуцеллус, который гомологичен мегаспорангию. У цветковых растений в нуцеллусе, вблизи верхушки (в области микропиле), из клеток археспория обособляется одна материнская клетка (мегаспора), имеющая, как и все растение, диплоидный набор хромосом. После мейотического деления этой клетки образуются 4 гаплоидные мегаспоры, располагающиеся, как правило, линейно - от микропиле к халазе. Таким образом, происходит мегаспорогенез.
После этого начинается мегагаметогенез, т.е. формирование женского гаметофита - зародышевого мешка. Из 4 образовавшихся мегаспор 3 дегенерируют и рассасываются, а 1 мегаспора, обращен- ная ближе к халазе, начинает делиться, и из нее развивается женский
гаметофит - зародышевый мешок. При этом ядро мегаспоры претерпевает 3 последовательных митотических деления (эндомитоз), в результате чего образуется 8 моноплоидных ядер. После 1-го деления мегаспоры образуются 2 ядра, которые расходятся к полюсам удлиняющейся мегаспоры. Далее каждое из этих ядер синхронно делится еще 2 раза; в результате у каждого полюса возникают по 4 ядра. На этой 8-ядерной стадии процесс деления ядра женского гаметофита заканчивается. При этом мегаспора разрастается, оттесняя ткань ну- целлуса к интегументам. От каждой из 2 четверок ядер в центральную часть зародышевого мешка отходит по 1 ядру. Эти так называемые полярные ядра сближаются в центральной части и, сливаясь, образуют диплоидное вторичное ядро. В дальнейшем 7 ядер зародышевого мешка претерпевают фазу цитоплазматического деления, т.е. каждое ядро с участком цитоплазмы окружается плазматической мембраной; следовательно, образуется 7 отдельных клеток (рис. 4.9, 4.10).
У микропилярного полюса гаметофита три клетки формируют яйцевой аппарат, одна из которых, наиболее крупная, называется яйце-
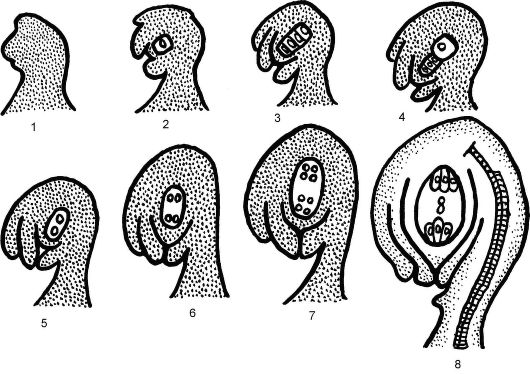
Рис. 4.9. Схема формирования зародышевого мешка: 1-4 - развитие нуцеллуса, обособление и мейоз клетки археспория, отмирание 3 мегаспор; 5-8 - развитие женского гаметофита - зародышевого мешка из оставшейся мегаспоры; 9 - 8-ядерный зародышевый мешок
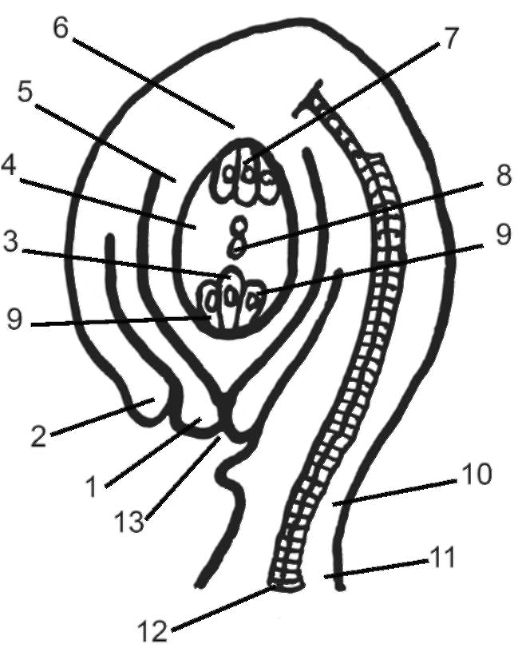
Рис. 4.10. Строение семязачатка: 1 - внутренний; 2 - наружный интегументы; 3 - яйцеклетка; 4 - зародышевый мешок; 5 - нуцеллус; 6 - халаза; 7 - антиподы; 8 - два полярных ядра; 9 - синергиды; 10 - фуникулюс; 11 - пла- цента; 12 - проводящий пучок; 13 - пыльцевход (микропиле)
клеткой, а две другие - вспомогательными клетками-синергидами. 3 клетки халазального полюса получили название антипод. В централь- ной части зародышевого мешка располагается клетка с диплоидным ядром. Так возникает женский гаметофит, который сильно редуцирован даже по сравнению с гаметофитом голосеменных и полностью лишен архегониев.
Опыление
Для образования зародыша должны произойти опыление и оплодотворение. Опыление - это процесс переноса пыльцы с тычинки на рыльце пестика. Опыление впервые появляется у голосеменных растений, но наибольшего совершенства оно достигает у покрытосеменных.
Различают 2 типа опыления: самоопыление и перекрестное опыление. При самоопылении на пестик цветка попадает пыльца того же растения; если перенос пыльцы осуществляется между цветками разных особей, происходит перекрестное опыление. Считается, что оно свойственно 90% растений. Перекрестное опыление обусловливает высокий уровень гетерозиготности популяций. Это создает большие возможности для естественного отбора. Строгое самоопыление встречается относительно редко (например, у гороха) и может вести к расщеплению вида на ряд чистых линий, т.е. делает популяции гомозиготными.
Для эволюционного процесса оптимальным считается сочетание самоопыления и перекрестного опыления, что чаще и происходит в природе. Одна из форм, ограничивающих самоопыление, - дву- домность, т.е. на одних растениях развиваются только мужские (тычиночные) цветки, а на других - женские (пестичные), имеющие только гинецей. Однодомные растения имеют цветки, содержащие и андроцей, и гинецей. Другая форма, ограничивающая самоопыление, - полная и физиологическая несовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи.
Выделяют 2 типа перекрестного опыления - биотическое и абиотическое. Биотическое опыление осуществляется животными, абиотическое - с помощью неживых факторов внешней среды (ветер, вода у водных растений).
Пыльца, попав на рыльце пестика, начинает прорастать. Из вегетативной клетки образуется пыльцевая трубка, а из генеративной - 2 спермия. Пыльцевая трубка проникает в зародышевый мешок и, достигнув яйцеклетки, лопается, что обеспечивает проникновение в него спермиев. Один спермий копулирует с яйцеклеткой, образуя зиготу, дающую начало зародышу. Второй спермий сливается с вторичным диплоидным ядром, располагающимся в центре зародышевого мешка, что приводит к образованию триплоидного ядра. В результате формируется триплоидная клетка, развивающаяся в специальную питательную ткань - эндосперм (3n) (от греч. endon - внутри и sperma - семя). Таким образом, происходит двойное оплодотворение, свойственное только покрытосеменным растениям. Оно впервые описано в 1898 г. выдающимся русским цитологом и эмбриологом С.Г. Навашиным. Прочие клетки зародышевого мешка - антиподы и синергиды - разрушаются.
Биологический смысл двойного оплодотворения состоит в том, что триплоидный эндосперм развивается лишь в случае оплодотворе- ния, чем достигается существенная экономия энергетических и пластических ресурсов в отличие от голосеменных растений, у которых образование эндосперма не связано с оплодотворением.
У покрытосеменных растений эндосперм называется «вторичным», или «белком». Только у покрытосеменных растений зародыш (спорофаза) начинает свое развитие самостоятельно за счет трипло-
идной фазы. У всех предшествующих групп (голосеменные и др.) зародыш развивается за счет гаметофазы.
Таким образом, из составных частей цветка образуются:
Из оплодотворенной яйцеклетки - зародыш (2n);
Из диплоидного ядра - эндосперм (3n);
. из интегументов семязачатка - семенная кожура семени (2n);
Из нуцеллуса - перисперм семени (2n);
Из стенок завязи, часто с участием других элементов цветка (чашечки, цветоложа), - стенка плода (перикарпий). Перикарпий имеет 3 слоя: наружный - экзокарпий, средний - мезокарпий и внутренний - эндокарпий.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис, при котором семена у апомиктичных растений образуются без оплодотворения из половой (партено- генез) или вегетативной (аногамия) клетки. При этом не происходит обмена генетической информацией, и поэтому все апомиктические особи имеют одинаковую генетическую и соматическую конституцию. Ярким примером растения с апомиктическим образованием семян по типу партеногенеза является одуванчик, отличающийся высокой жизнеспособностью. Нередко, особенно у многолетних корневищных растений, преобладает вегетативное размножение, а семенное - подавлено.
Соцветия
Соцветия - это побег или система специализированных побегов, несущих цветки. Соцветия свойственны громадному большинству цветковых растений. Соцветие имеет главную ось (ось соцветия) и боковые. Последние могут ветвиться или быть неразветвленными и нести цветки. На осях соцветий имеются узлы и междоузлия. На узлах соцветия располагаются прицветники, а на узлах цветоножки - прицветнички.
Биологический смысл возникновения соцветия - в возрастающей вероятности опыления цветков. Несомненно, что соцветия более за- метны среди зелени листьев, нежели единичные цветки, и насекомое за единицу времени посетит гораздо больше цветков, если они собраны в соцветия. Обычно соцветия группируются близ верхней части растения на концах ветвей.
Классификация соцветий достаточно сложна, но в справочниках и определителях по систематике растений учитывают 2 признака: характер ветвления и способ нарастания.
В зависимости от степени разветвления осей соцветия делят на простые и сложные. У простых соцветий на главной оси располагаются одиночные цветки (черемуха, подорожник, подсолнечник); у сложных - не одиночные цветки, а боковые оси (донник лекарственный, сирень, пшеница, мятлик). У одних растений апикальные меристемы расходуются на формирование верхушечного цветка, и в этом случае соцветия относят к закрытым, или определенным. У закрытых соцветий (симподиальных) верхушечные цветки обычно раскрываются раньше нижележащих боковых, поэтому их называют верхо цветными. У других растений апикальные меристемы остаются в вегетативном состоянии, и такие соцветия называют открытыми, или неопределен ными. У открытых (моноподиальных) соцветий цветки распускаются последовательно снизу вверх, поэтому их называют бокоцветными.
Существует несколько типов ботрических (от греч. botris - кисть) соцветий (рис. 4.11).
Простые ботрические соцветия - это моноподиальные соцветия, характеризующиеся длиной и формой главной оси, а также наличием или отсутствием цветоножек и прицветников.
Кисть (основной вариант простых соцветий) - ось тонкая, с цветками на цветоножках более или менее одинаковой длины, которые поочередно прикрепляются к главней оси соцветия. Кисти бывают односторонние (ландыш) и двусторонние (черемуха).
Щиток - соцветие, сходное с кистью, но у щитка нижние цвето- ножки длиннее верхних, поэтому его цветки расположены на одном уровне и в одной плоскости (рябина, боярышник).
Зонтик - производное кисти с сильно укороченной тонкой осью и цветками на цветоножках одинаковой длины (вишня, чистотел, примула).
Головка - видоизмененный зонтик, у которого главная ось раз- растается в виде головки, а на ней располагаются цветки на укороченных цветоножках (клевер, люцерна).
Колос - производное кисти, отличающееся от нее отсутствием цветоножек (подорожник).
Сережка - повислый колос, т.е. колос с мягкой осью, несущий однополые цветки; после цветения соцветие обычно целиком опадает (ива, тополь).
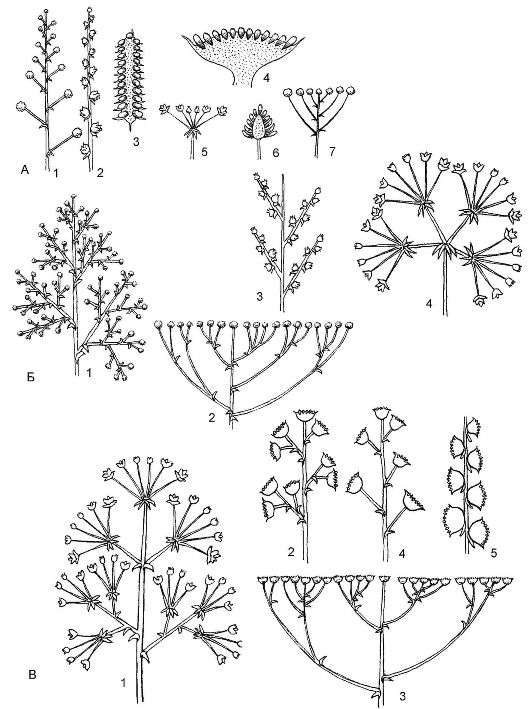
Рис. 4.11. Типы ботрических соцветий: А - простые: 1 - кисть; 2 - колос; 3 - початок; 4 - корзинка; 5 - простой зонтик; 6 - головка; 7 - щиток;
Б - сложные: 1 - метелка; 2 - сложный щиток; 3 - двойной колос; 4 - сложный зонтик;
В - типы агрегатных соцветий: 1 - метелка зонтиков; 2 - метелка корзинок; 3 - щиток корзинок; 4 - кисть корзинок; 5 - колос корзинок
Початок - разновидность колоса, но с толстой мясистой осью (кукуруза). Часто початок окружен листом разной формы и окраски, который называют покрывалом или крылом.
Корзинка - главная ось соцветия разрастается в виде чаши или ложа, на котором располагаются сидячие цветки, окруженные со всех сторон (часто в 2 ряда) видоизмененными присоцветными листьями - листовой оберткой (подсолнечник, ромашка). Поскольку корзинка - соцветие моноподиальное, распускание цветков в ней идет центростремительно - от периферии к центру.
Сложные ботрические соцветия могут быть как открытыми, так и закрытыми.
Сложная кисть (метелка) - соцветие, у которого на удлиненной моноподиальной главной оси располагаются ботрические парциальные (или частные) соцветия, являющиеся простыми кистями. В зависимости от степени ветвления кисть может быть двойная - соцветие, у которого на главной оси располагаются простые кисти (донник, вероника простертая); тройная - имеющая оси 3-го порядка (хрен, алоэ крупноплодное). К сложным метелкам относятся составные (агрегатные) соцветия. Они характеризуются тем, что тип нарастания главной оси отличается от такового у парциальных соцветий - например, метелка корзинок (полынь обыкновенная), кисть зонтиков (плющ обыкновенный), колос корзинок (сушеница лесная).
Сложный колос - соцветие, у которого на удлиненной главной оси располагаются простые колосья, называемые колосками (пшеница, рожь, ячмень); морфологически близко к двойной кисти.
Сложный зонтик - соцветие с укороченной главной осью, на которой расположена розетка верховых листьев (прицветников), называемая общей оберткой. Из пазух листьев выходят оси, завершающиеся парциальными соцветиями - простыми зонтиками, которые называют зонтичками. Цветки в зонтичках часто имеют выраженные прицветники, составляющие оберточки. Сложные зонтики свойственны многим растениям семейства Сельдерейные (морковь, пе- трушка, укроп, сельдерей). Это открытые соцветия, поэтому цветки в них распускаются центростремительно.
Цимозные (симподиальные соцветия) (от греч. kyma - волна) получили свое название по особому способу зацветания. Их соцветия закрытые, главная ось у них не выражена, а парциальные соцветия имеют симподиальное ветвление. Цветки возникают в пазухах прицветных листьев на осях предыдущих порядков, т.е. распускание цветков происходит центробежно. В зависимости от числа боковых
осей цимоиды разделяют на три основных типа: монохазий, дихазий, плейохазий (рис. 4.12).
Монохазий (однолучевик) - такое цимозное соцветие, у которого каждая материнская ось несет 1 дочернюю, формирующуюся под цветком, завершающим ось предыдущего порядка. В зависимости от направления осей различают 2 типа монохазиев: завиток и извилина.
Завиток (улитка) - от оси 1-го порядка отходит ось 2-го порядка, от нее - 3-го порядка и т.д., но при этом оси отходят только в одном направлении, как бы закручиваясь в улитку или завиток (незабудка и другие представители семейства Бурачниковые).
Извилина - от оси первого порядка отходит ось второго порядка, от второго - третьего и т.д., то в одну, то в другую сторону (лютик едкий).
Дихазий (двулучевик) - соцветие, у которого под верхушечным цветком главной оси развиваются 2 парциальных соцветия, при этом
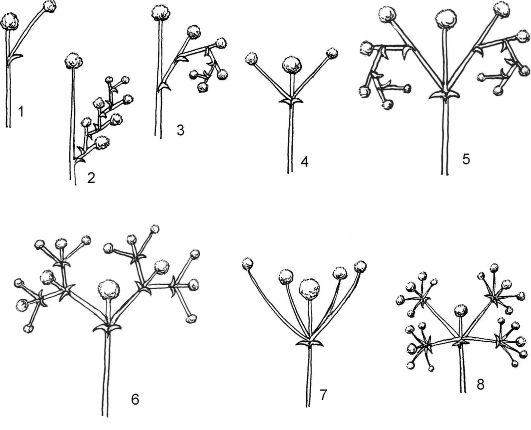
Рис. 4.12. Типы цимоидных соцветий: монохазии: 1 - «элементарный» монохазий; 2 - извилина; 3 - завиток; дихазии: 4 - простой; 5 - двойной, или двойной завиток; 6 - тройной; плейохазии: 7 - простой; 8 - двойной
каждая ось предыдущего порядка несет 2 оси следующего порядка. К дихазиям относятся развилина и двойной завиток.
Развилина - под цветком главной оси образуются супротивно рас- положенные ветви (оси), заканчивающиеся цветками. В дальнейшем каждая из этих осей также образует 2 супротивно расположенные оси следующего порядка (гвоздика).
Двойной завиток - под цветком главной оси супротивно образуются 2 простых завитка (картофель).
Плейохазий (многолучевик) - соцветие, у которого каждую ось предыдущего порядка сменяют более 2 осей следующего порядка (молочай, родиола).
Цветение. Однолетние растения зацветают не ранее чем через 20- 30 дней после посева. Все однолетние (рожь, пшеница) и двулетние (редька, капуста, морковь) растения цветут и плодоносят один раз в жизни, после чего погибают, поэтому их называют монокарпическими (или монокарпиками). Многолетние травы зацветают на 2-й год жизни, а многие деревья - через десятки лет. У многолетних растений, как правило, цветение и плодоношение многократно повторяются в течение жизни, поэтому их называют поликарпическими (или поли- карпиками). Лишь немногие многолетние растения являются монокарпиками, т.е. после первого цветения и плодоношения отмирают (некоторые агавы, бамбуки, пальмы, ферулы).
Вопросы к коллоквиуму по теме
для студентов-биологов II курса ДО
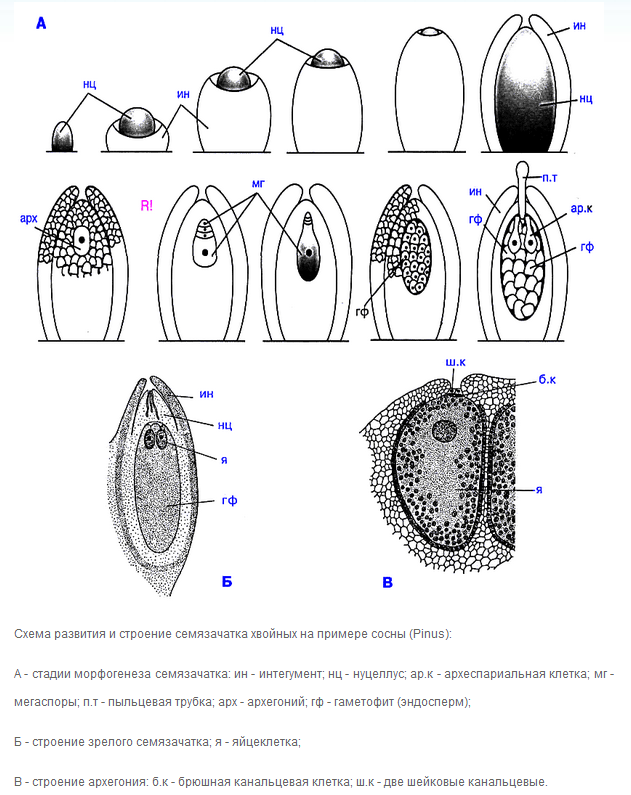
Что такое семязачаток? Перечислите его составные части у Голосеменных.
Семязача́ток , или семяпо́чка (лат. ovulum ) - образование у семенных растений, из которого (обычно после оплодотворения) развивается семя. Представляет собой женский спорангий (мегаспорангий) семенных растений. У голосеменных - на поверхности семенных чешуек в женских шишках, расположен открыто в пазухе мегаспорофилла. В центральной части семяпочки (нуцеллусе) формируются в результате мейоза материнской клетки спор четыре мегаспоры, затем три из них гибнут, а из одной мегаспоры формируется женский гаметофит. У голосеменных его иногда называют эндосперм, так как в зрелом семени в нём запасаются питательные вещества.
С. состоит из нуцеллуса (центр. части, содержащей мегаспороцит) , одного или двух покровов (интегументов) и семяножки (фуникулуса) . На верхушке С. покровы обычно не смыкаются, остаётся узкое отверстие - микропиле.
Что такое нуцеллюс? Как происходит закладка и развитие семязачатка?
Нуц е ллус (от лат. nucella - орешек), центральная часть (ядро) семяпочки семенных растений; гомологиченмегаспорангию папоротникообразных. Включает питательную ткань и зародышевый мешочек, покрытый оболочкой, с маленьким отверстием, называемым микропилем.
Теории происхождения интегумента семязачатка.
Теломная
интегумент представляет собой результат срастания периферических изначально вегетативных теломов вокруг одного спороносного. Эта теория согласуется с палеоботаническими находками, которые относятся к праголосеменным и вымершим голосеменным - семенным папоротникам.
Синангиальная
Согласно этой гипотезе, выдвинутой впервые английским палеоботаником Маргаритой Бенсон (1908), интегумент представляет собой кольцо подвергшихся стерилизации, сросшихся и слившихся между собой спорангиев, окружающих центральный функционирующий мегаспорангий, а микропиле соответствует первоначальному промежутку между верхушками спорангиев. Другими словами, семязачаток является фактически синангием, у которого все спорангии, кроме одного, стерилизовались и образовали покров (интегумент) единственного, фертильного спорангия. Хорошим подтверждением «синангиальной» гипотезы являются примитивные семязачатки семенных папоротников, сохранившие часто весьма ясные следы своего синангиального происхождения. Семязачатки ряда семенных папоротников обладали сегментированными интегументами с проводящим пучком в каждом сегменте (камере).
Микропилярный и халазальный полюса семязачатка
В семенном зачатке халаза находится напротив микропиле, открывая интегумент. Халаза - это ткань где соединяются интегумент и нуцеллус.
Два полюса зародыша соответствуют корневому и побеговому полюсам.
В зародыше: гипокотиль и семядоли
Особенности семязачатка как видоизменённого мегаспорангия семенных растений в сравнении с мегаспорангием высших споровых.
Семя – новый тип диаспор.
Процесс мегаспорогенеза
На чешуях женских шишек находятся 2 семязачатка. Семязачаток состоит из нуцеллуса (ядра) и интегумента (покров). Наверху остается небольшое отверстие – микропиле (пыльцевход). Поздней весной чешуи женской шишки раскрываются и пыльца попадает через микропиле внутрь на нуцеллус. После этого семенные чешуи сжимаются, образуя защиту семязачатков. Во время опыления в пылинке нет еще мужских гамет, а в семязачатке не развит еще заросток с архегониями. Через месяц после опыления одна из клеток нуцеллуса начинает делиться мейозом. В результате образуются 4 гаплоидные (n) клетки – мегаспоры. 3 отмирают, а 1 превращается в заросток (n). Женский гаметофит представляет собой бесцветный многоклеточный таллом. ^ В ткани много запасающих веществ. Через 14-15 месяцев после опыления на гаметофите образуются два архегония. Архегоний состоит из крупной яйцеклетки с большим ядром, над которой лежит рано исчезающая брюшная канальцевая клетка, и шейки из восьми мелких клеток. В это время пыльцевая трубка очень медленно растет внутрь нуцеллуса. После того, как пыльцевая трубка с двумя спермиями достигнет архегонии, один из спермиев сливается с яйцеклеткой, другой гибнет. Из зиготы развивается зародыш. Семязачаток превращается в семя. Вегетативное тело женского гаметофита становится первичным эндоспермом (n). Из нуцеусса образуется пленчатая кожура, из интегумента – деревянистая. Таким образом в семени можно выделить части 3 поколений:
Деревянистая и пленчатая кожура – старый спорофит (2n)
Эндосперм – гаметофит (n)
Зародыш – новый спорофит (2n)
Типичный вариант развития женского гаметофита Голосеменных
Каким структурам гомологичны пыльники и пыльцевые зёрна Голосеменных?
Пыльник – микроспорангий, пылинка – пыльцевое зерно
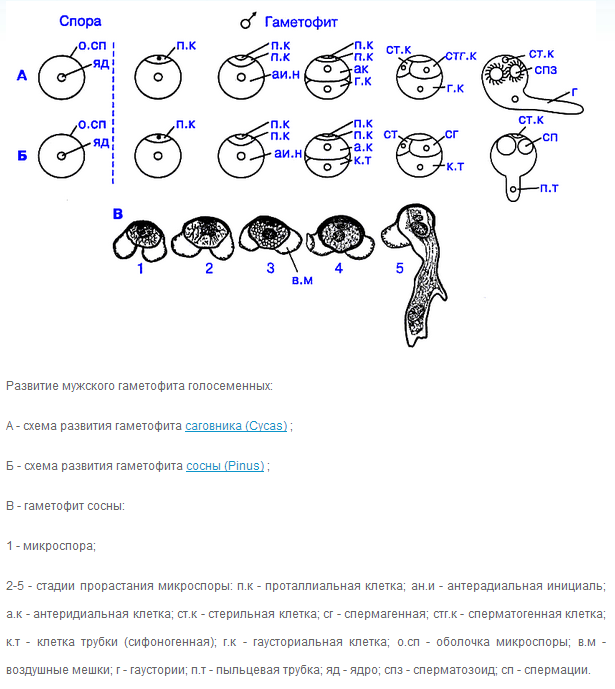
10. 2 варианта строения зрелого мужского гаметофита, его клеточные элементы
11. Дальнейшая судьба мужского гаметофита Голосеменных.
12. Всегда ли пыльцевое зерно покидает пыльник зрелым? Какой вариант более прогрессивен?
13. Опыление (определение)
У растений, перенос пыльцы с пыльников на рыльце пестика (у цветковых растений) или на семяпочку (у голосеменных). После О. из пылинки развивается пыльцевая трубка, к-рая растёт в сторону завязи и доставляет муж. половые клетки - спермин - к яйцеклетке, находящейся в семяпочке, где и происходит оплодотворение и развитие зародыша.
14. Оплодотворение типа саговниковых, его особенности
После опыления семязачатки начинают увеличиваться и скоро достигают размеров семени, хотя оплодотворение в них еще не произошло. Этот период, от опыления до оплодотворения, весьма длителен и занимает обычно полгода (так, у саговника завитого опыление происходит в декабре - январе, а оплодотворение - в мае - июне).

Микроспоры, попавшие с капелькой опылительной жидкости в пыльцевую камеру, прорастают. Экзина при этом лопается, и через разрыв прорастает, растягивая интину, клеткагаустория (рис. 168, 13). Она внедряется в стенку пыльцевой камеры и высасывает из ткани нуцеллуса питательные вещества (рис. 168, 13). Генеративная клетка в это время делится на две, и одна из образовавшихся клеток - сперматогенная - начинает усиленно расти. В ней и формируются, не сразу, а спустя несколько месяцев, мужские гаметы - сперматозоиды (рис. 168, 15).
К моменту оплодотворения разросшаяся сперматогенная клетка оказывается в непосредственной близости от входа в архегонии. Освободившимся из нее сперматозоидам остается «проплыть» в жидкости, которая излилась вместе с ними из сперматогенной клетки, лишь небольшое расстояние до архегония, в котором происходит слияние содержимого сперматозоида с яйцеклеткой (рис. 168, 17).
Таким образом, у саговниковых в едином процессе соединяются два механизма, один из которых - образование подвижного сперматозоида - свойствен далеким оплодотворявшимся с помощью воды предкам, а второй - формирование пыльцевой трубки (разрастающаяся сперматогенная клетка) - типичен для всех остальных стоящих выше на «эволюционной лестнице» семенных растений.
15. Сифоногамия
(оплодотворение типа сосновых) – оплодотворение с помощью пыльцевой трубки; половой процесс, протекающий внутри семязачатков и не зависит от наличия влаги. Функцию доставки мужских гамет осуществляют особые клетки.
16. Чем принципиально различаются данные варианты оплодотворения у Голосеменных?
17. Какие клетки (или их особенности) в первом и во втором случае можно назвать атавистическими?
Семя развивается на поверхности семенной чешуи. Оно представляет собой многоклеточную структуру, объединяющую запасающую ткань - эндосперм, зародыш и специальный защитный покров (семенную кожуру). До оплодотворения в центральной части семязачатка имеется нуцеллус, который постепенно вытесняется эндоспермом. Эндосперм гаплоидный и образуется из тканей женского гаметофита.
У саговников и гинкго наружный слой семенной кожуры (саркотеста ) мягкий и мясистый, средний слой (склеротеста ) твёрдый, а внутренний слой (эндотеста) к моменту созревания семени плёнчатый. Семена распространяются различными животными, которые поедают саркотесту, не повреждая склеротесты.
У тиса и подокарпуса семена окружены мясистым ариллусом - сильно видоизменённой чешуёй женской шишки. Сочный и ярко окрашенный ариллус привлекает птиц, которые распространяют семена этих хвойных. Ариллусы многих видов подокарпуса съедобны и для человека.
19. Из каких структур семязачатка развиваются соответствующие части семени?
20. Структуры каких стадий жизненного цикла включает семя?
22. Сравните строение семени гинкго и сосны. В чём проявляются признаки примитивности первого?
Характерный для сосен развитый подвесок ко времени полного развития зародыша дегенерирует. Семя сосны состоит из зародыша, семенной кожуры и мегагаметофита, представляющего собой запас питательных веществ.
У саговников и гинкго наружный слой семенной кожуры (саркотеста) мягкий и мясистый, средний слой (склеротеста) твёрдый, а внутренний слой (эндотеста) к моменту созревания семени плёнчатый.
|
семязачатки гинкго платиспермические | ||||
|
Дополнительные структуры объединяют под термином ангиоспермизация | ||||
|
Пыльцевая камера унитегмальна | ||||
|
Оплодотворение после опадения | ||||
23. Что такое первичный эндосперм семени? С чем связано его название?
Первичный эндосперм голосеменных образуется ДО ОПЛОДОТВОРЕНИЯ из мегаспоры и соответствует женскому гаметофиту. Клетки эндосперма голосеменных первоначально гаплоидные, затем в результате слияния ядер становятся полиплоидными.
Втори́чный эндоспе́рм - ткань, образующаяся в семенах большинства цветковых растений ВО ВРЕМЯ ОПЛОДОТВОРЕНИЯ.
24. Эволюционные преимущества семенного размножения.
Семена более жизнеспособно, благодаря животным, ветру, воде могут переноситься на большие расстояния. Есть запас питательных веществ, зародыш защищен оболочками семени. Размножение не связано с водой.
Строение семяпочки голосеменных . Женская шишка состоит из довольно толстого стержня - оси, к которой спирально прикрепляются семенные чешуи. На каждой семенной чешуе, у ее основания, расположены по два довольно крупных семязачатка (мегаспорангия). Под каждой семенной чешуей расположена другая маленькая чешуя, которая называется кроющей. Семязачаток снаружи имеет покров, или интегумент. Под покровом находится овальное тело - нуцеллус. С наружного конца семязачатка имеется два небольших выроста. Между ними расположен пыльцевход (микропиле), который представляет собой тонкое отверстие, идущее внутрь семязачатка.
Одна из клеток нуцеллуса обособляется и делится редукционным путем, в результате чего образуется четыре гаплоидные клетки - мегаспоры, расположенные друг над другом. Три верхние клетки отмирают, а четвертая (нижняя) увеличивается в размерах, делится и образует женский заросток, или женский гаметофит, который является также гаплоидным эндоспермом. В верхней части заростка расположено обычно по два сильно редуцированных архегония с очень крупными яйцеклетками.
Большое количество пыльцы гибнет при переносе ветром, и лишь небольшая ее часть попадает на семязачатки женских шишек, где она прорастает. Пыльцевое зерно через пыльцевход вовлекается внутрь семязачатка. Вегетативная клетка принимает участие в образовании пыльцевой трубочки. Через ткани нуцеллуса пыльцевая трубочка проникает к архегонию. По пыльцевой трубочке проходит антеридиальная клетка, которая еще в пыльцевой трубочке делится на два спермия. В процессе оплодотворения происходит слияние яйцеклетки с одним из спермиев (другой спермий погибает) и образуется диплоидная зигота, из которой формируется зародыш; он состоит из зачаточного корешка, стебелька, семядолей и почечки.
После оплодотворения из семязачатка образуется семя, покрытое твердой кожурой, развивающейся из покрова (интегумента). Остатки нуцеллуса сохраняются в виде тонкой пленки, окружающей эндосперм, который занимает центральную часть семени и является питательной тканью; в нем расположен зародыш семени, который является молодым спорофитом. В период формирования семян женские шишки сильно разрастаются. Ко времени созревания семян семенные чешуи подсыхают и окрашиваются в коричневый цвет. Споролистики в созревшей шишке слегка расходятся друг от друга и семена свободно могут высыпаться из шишки. Семена сосны имеют прозрачный крылатый вырост, с помощью которого разносятся ветром. В благоприятных условиях семена прорастают и образуют крупные растения (спорофиты).
Строение семязачатка (семяпочки) покрытосеменных . Семязачаток снаружи бывает покрыт покровами, или интегументами. Семязачаток может иметь один или два покрова. Один из них наружный, другой - внутренний. Под покровами находится особая паренхимная ткань. Она составляет центральную часть семязачатка и называется нуцеллусом или ядром семязачатка. Нуцеллус снаружи защищен покровами семязачатка. В верхней части семязачатка покровы не соединяются и между ними остается небольшое отверстие, которое называется пыльцевходом, или семявходом.. Во внутренней части семязачатка, в нуцеллусе, находится зародышевый мешок, в котором происходит процесс оплодотворения.
Зародышевый мешок представляет собой женский гаметофит, т. е. половое поколение. Развитие зародышевого мешка происходит из одной клетки нуцеллуса. Ядро этой клетки делится мейотичееким делением, в результате чего возникает четыре клетки - тетрада. Одна из четырех клеток развивается и превращается в зародышевый мешок. Во вполне сформировавшемся зародышевом мешке имеется 7 клеток. В центре зародышевого мешка находится центральное, или вторичное, ядро, которое образовалось от слияния двух ядер. На каждом из противоположных полюсов расположено по о клетки. Из 3 клеток, расположенных около семявхода, или пыльцевхода, состоит так называемый яйцевой аппарат.
В яйцевой аппарат входят яйцеклетка и две вспомогательные клетки, или синергиды. На противоположном конце зародышевого мешка, т. е. напротив яйцевого аппарата, располагаются еще 3 клетки (антиподы). Центральное ядрозародышевого мешка, или вторичное ядро, произошло от слияния двух ядер, поэтому оно имеет двойное, или диплоидное, число хромосом (2n). Остальные клетки зародышевого мешка (в том числе и яйцеклетка) имеют одинарный набор хромосом, т. е. гаплоидный (n). В таком виде зародышевый мешок подготовлен к процессу оплодотворения. Такое строение зародышевого мешка характерно для покрытосеменных растений. Основание нуцеллуса и покровов семязачатка расположено на противоположной стороне от пыльцевхода. Это место называется халазой. Семязачатки обычно имеют овальнояйцевидную форму. По своему положению по отношению к семяножке различают семязачатки прямые, обратные и согнутые.
Процессу оплодотворения у цветковых растений предшествует процесс опыления, в результате которого пыльца тем или иным способом попадает на рыльце пестика. Между этими двумя процессами, т. е. опылением и оплодотворением, проходит довольно много времени. Процесс прорастания пыльцы заключается в следующем: все содержимое пылинки, одетое оболочкой интиной, начинает выпячиваться через поры в наружной оболочке экзине, в результате чего постепенно образуется пыльцевая трубочка, которая внедряется в ткань рыльца пестика.Генеративное ядро, находящееся в пылинке, делится на два спермия, но чаще это разделение происходит в пыльцевой трубочке. В семязачатке пыльцевая трубочка через ткани нуцеллуса подходит к зародышевому мешку, врастает в его ткань, разрушая при этом одну из клеток яйцевого аппарата, т. е. одну из двух синергид. В зародышевом мешке конец пыльцевой трубочки лопается и оба спермин попадают внутрь зародышевого мешка, где происходит процесс оплодотворения. Один из двух спермиев сливается с женской половой клеткой ~ яйцеклеткой, другой - со вторичным ядром зародышевого мешка. Таким образом, оба спермия принимают участие в оплодотворении. Такой тип оплодотворения является особенностью покрытосеменных растений и называется двойным оплодотворением. Двойное оплодотворение было открыто русским ученым С. Г Навашиным (1898).
При оплодотворении яйцеклетки одним из спермиев образуется зигота, которая имеет двойной набор хромосом. В дальнейшем из оплодотворенной яйцеклетки развивается зародыш семени. При оплодотворении центрального или вторичного ядра зародышевого мешка другим сиермием возникает клетка с триплоидным набором хромосом в ядре, из которой впоследствии развивается питательная ткань семени - эндосперм. После оплодотворения вторичное ядро находится некоторое время в стадии покоя, затем начинает активно 1 делиться и постепенно заполняет почти всю полость зародышевого мешка, превращаясь в питательную ткань.
После процесса оплодотворения наступают большие преобразования в зародышевом мешке и во всем семязачатке в целом. Преобразования также захватывают всю завязь пестика. Весь семязачаток после оплодотворения превращается в семя. Из покровов семязачатка образуется кожура семени. Завязь превращается в плод. Стенки завязи образуют стенки плода, которые называются околоплодником. Иногда в образовании плода принимают участие также другие части цветка.
После оплодотворения семязачаток называют уже семенем , а завязь - плодом .
Зигота растет путем многократных митотических делений и превращается в многоклеточный зародыш, состоящий из первичного побега, или плюмулы, первичного корешка и одной (у однодольных) или двух (у двудольных) семядолей . Семядоли имеют более простую структуру, чем первые настоящие листья; иногда они наполнены питательными веществами и служат запасающей тканью, как, например, у гороха и у конских бобов (Vicia faba ). Плюмула состоит из стебля, первой пары настоящих листьев и терминальной почки. Триплоидное ядро первичного эндосперма многократно делится (путем митоза), образуя эндосперм - скопление триплоидных ядер, отделенных друг от друга тонкими клеточными стенками. У некоторых растений, например у кукурузы (Zea mays ), эндосперм сохраняет роль запасающей ткани.
Если запасающей тканью служат семядоли, то они растут за счет эндосперма, который может при этом совершенно исчезнуть. У некоторых семян запасы питательных веществ содержатся и в эндосперме, и в семядолях.
В развивающемся семени рост зародыша, а иногда и эндосперма происходит в зародышевом мешке. По мере продолжения роста окружающий их нуцеллус дезорганизуется и разрушается, доставляя необходимые для роста питательные вещества. В дальнейшем снабжение питательными веществами обеспечивает проводящий пучок фуникулуса (ножки семязачатка), подобно тому как развивающийся зародыш млекопитающего получает питание от материнского организма через плаценту. Как уже отмечалось, место прикрепления фуникулуса к стенке завязи тоже называется плацентой.
Микропиле сохраняется в виде маленькой поры в тесте (семенной кожуре), через которую поступают кислород и вода при прорастании семени. Теста - это тонкий, но прочный защитный слой, образующийся из интегументов. На завершающих стадиях созревания семени содержание в нем воды снижается от обычного для растительных тканей - примерно 90% (по массе) - до 10-15%. Это сопровождается заметным снижением метаболической активности и представляет собой важный шаг к переходу семени в состояние покоя.
По мере развития семени завязь превращается в зрелый плод и стенка ее получает название перикарпия . Происходящие при этом изменения у разных видов различны, но обычно направлены на то, чтобы плод мог защищать семена и способствовать их распространению (см. разд. 20.2.8).
Остальные части цветка увядают, отмирают и сбрасываются, подобно листьям у листопадных растений. В некоторых случаях такие части, как цветоложе, столбик или чашелистики сохраняются и участвуют в распространении семян; цветоложе в таких случаях, например у земляники (Fragaria ), получает название ложного плода .
Гормональная регуляция развития плода рассмотрена в разд. 15.3.5.
Некоторые изменения, происходящие после оплодотворения, суммированы в табл. 20.3.
Некоторые стадии развития зародыша показаны на рис. 20.27.