Ang germinal stage ng isang seed plant, na nabuo sa proseso ng sekswal na pagpaparami at nagsisilbi para sa pag-aayos. Sa loob ng buto ay isang embryo, na binubuo ng isang embryonic root, isang tangkay at isa o dalawang dahon, o cotyledon. Ayon sa bilang ng mga cotyledon, ang mga halaman ay nahahati sa dicots at monocots. Sa ilang mga species, tulad ng mga orchid, ang mga indibidwal na bahagi ng embryo ay hindi naiiba at nagsisimulang mabuo mula sa ilang mga cell kaagad pagkatapos ng pagtubo. Ang isang tipikal na buto ay naglalaman ng suplay ng mga sustansya para sa embryo, na kailangang tumubo nang ilang panahon nang walang liwanag na kailangan. Ang reserbang ito ay maaaring sakupin ang karamihan sa mga buto, at kung minsan ay matatagpuan sa loob ng embryo mismo - sa mga cotyledon nito (halimbawa, sa mga gisantes o beans); pagkatapos ang mga ito ay malaki, mataba at tinutukoy ang pangkalahatang hugis ng buto. Kapag tumubo ang buto, maaari silang kunin sa lupa sa isang pahabang tangkay at maging unang mga dahon ng photosynthetic. batang halaman. Sa mga monocots (halimbawa, trigo at mais), ang supply ng pagkain ay ang tinatawag na. endosperm - palaging hiwalay sa embryo. Ang ground endosperm ng mga pananim na butil ay isang kilalang harina. Natakpan halamang binhi ang buto ay bubuo mula sa ovule - isang maliit na pampalapot sa panloob na dingding ng obaryo, i.e. ang ilalim ng pistil na matatagpuan sa gitna ng bulaklak. Ang obaryo ay maaaring maglaman ng isa hanggang ilang libong ovule. Ang bawat isa sa kanila ay naglalaman ng isang itlog. Kung, bilang isang resulta ng polinasyon, ito ay pinataba ng tamud na tumagos sa obaryo mula sa butil ng pollen, ang ovule ay bubuo sa isang buto. Lumalaki ito, at ang shell nito ay nagiging siksik at nagiging isang dalawang-layer na seed coat. Ang panloob na layer nito ay walang kulay, malansa at may kakayahang bumukol nang malakas, sumisipsip ng tubig. Ito ay magiging kapaki-pakinabang sa ibang pagkakataon, kapag ang lumalagong embryo ay kailangang masira ang seed coat. Ang panlabas na layer ay maaaring mamantika, malambot, mala-pelikula, matigas, mala-papel, at maging makahoy. Sa balat ng buto, ang tinatawag na. peklat - ang lugar kung saan ang buto ay konektado sa tangkay ng buto, na nakakabit nito sa magulang na organismo. Ang binhi ay ang batayan ng pagkakaroon ng modernong flora at fauna. Kung walang binhi, walang coniferous taiga, nangungulag na kagubatan, namumulaklak na parang, steppes, butil sa planeta, walang mga ibon at langgam, bubuyog at butterflies, mga tao at iba pang mga mammal. Ang lahat ng ito ay lumitaw lamang pagkatapos na ang mga halaman sa kurso ng ebolusyon ay may mga buto, sa loob kung saan ang buhay ay maaaring, nang hindi ipinapahayag ang sarili sa anumang paraan, ay mapangalagaan sa loob ng mga linggo, buwan at kahit na sa maraming taon. Miniature halamang mikrobyo sa buto ay nakakapaglakbay ng malalayong distansya; hindi siya nakatali sa lupa sa pamamagitan ng mga ugat, tulad ng kanyang mga magulang; hindi nangangailangan ng tubig o oxygen; naghihintay siya sa mga pakpak upang makarating sa isang angkop na lugar at maghintay para sa kanais-nais na mga kondisyon, upang simulan ang pag-unlad, na tinatawag na pagtubo ng binhi.
MGA URI NG BINHI. Mais - monocot namumulaklak na halaman, ang buto nito ay nasa loob ng prutas na tinatawag na caryopsis. Tulad ng lahat ng monocots, ang buto ay may isang cotyledon. Ang bulto ng butil ay puno ng endosperm - isang supply ng nutrients na ginagamit ng embryo ng halaman sa panahon ng pagtubo. - halaman ng gymnosperm. Sa bawat sukat ng babaeng cone nito, dalawang buto ang hayagang matatagpuan. Sa ilalim ng balat mayroon silang isang endosperm at isang embryo na may ilang mga cotyledon.
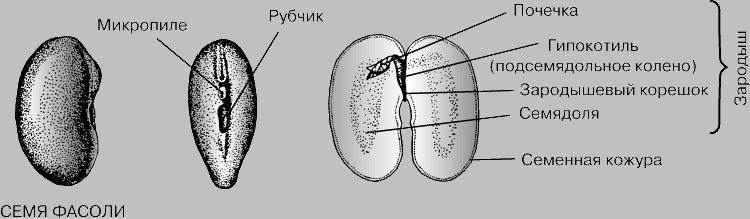
BEANS- isang dicotyledonous na namumulaklak na halaman, ang mga buto nito ay hinog sa loob ng beans. Walang endosperm sa loob ng buto, at ang buong supply ng nutrients na kailangan para sa pagbuo ng embryo ay naka-imbak sa dalawang malalaking mataba na cotyledon. Sa labas sa buto, ang isang peklat at isang micropyle ay maaaring makilala.
Ang ebolusyon ng mga buto. Sa daan-daang milyong taon, ang buhay sa Earth ay walang mga buto, tulad ng ginagawa nito nang wala sila ngayon sa dalawang-katlo ng ibabaw ng planeta na natatakpan ng tubig. Nagmula ang buhay sa dagat, at ang mga unang halaman na sumakop sa lupa ay wala pa ring binhi, ngunit ang hitsura lamang ng mga buto ang nagpapahintulot sa mga organismong photosynthetic na ganap na makabisado ang bagong tirahan na ito para sa kanila.
Ang mga unang halaman sa lupa. Sa malalaking organismo, ang unang pagtatangka na magkaroon ng panghahawakan sa lupa ay malamang na ginawa ng marine macrophytes - na napunta sa mga batong pinainit ng araw sa low tide. Nag-reproduce sila sa pamamagitan ng spores - mga single-celled na istruktura na dispersed ng magulang na organismo at may kakayahang umunlad sa isang bagong halaman. Ang mga spores ng algae ay napapalibutan ng manipis na mga shell, kaya hindi nila pinahihintulutan ang pagpapatayo. Sa ilalim ng tubig, ang gayong proteksyon ay sapat na. Ang mga spores doon ay kumakalat sa pamamagitan ng mga agos, at dahil ang temperatura ng tubig ay medyo mababago, hindi nila kailangang maghintay ng matagal para sa kanais-nais na mga kondisyon para sa pagtubo. Ang mga unang halaman sa lupa ay muling ginawa ng mga spore, ngunit ang sapilitan na pagbabago ng mga henerasyon ay naayos na sa kanilang ikot ng buhay. Ang prosesong sekswal na kasama dito ay tiniyak ang kumbinasyon ng mga namamana na katangian ng mga magulang, bilang isang resulta kung saan pinagsama ng mga supling ang mga pakinabang ng bawat isa sa kanila, nagiging mas malaki, mas matatag, at mas perpekto sa istraktura. Sa isang tiyak na yugto, ang gayong progresibong ebolusyon ay humantong sa paglitaw ng mga liverworts, mosses, club mosses, ferns at horsetails, na ganap na umalis sa mga reservoir sa lupa. Gayunpaman, hindi pa pinahintulutan ng mga spores na kumalat sila sa kabila ng mga marshy na lugar na may basa-basa at mainit na hangin.
Mga spore na halaman ng panahon ng Carboniferous. Sa yugtong ito ng pag-unlad ng Earth (mga 250 milyong taon na ang nakalilipas), ang mga higanteng anyo na may bahagyang lignified trunks ay lumitaw sa mga ferns at lycopsid. Hindi mas mababa sa kanila sa laki at, ang mga guwang na tangkay ay natatakpan ng berdeng bark na binasa sa silica. Saanman lumitaw ang mga halaman, sinusundan sila ng mga hayop, na pinagkadalubhasaan ang mga bagong uri ng tirahan. Sa mahalumigmig na takip-silim ng kagubatan ng karbon, mayroong maraming malalaking insekto (hanggang sa 30 cm ang haba), mga higanteng alupihan, mga gagamba at alakdan, mga amphibian na mukhang malalaking buwaya, at mga salamander. Mayroong mga tutubi na may wingspan na 74 cm at mga ipis na 10 cm ang haba. Ang mga tree ferns, club mosses at horsetails ay mayroong lahat ng mga katangian na kinakailangan para sa pamumuhay sa lupa, maliban sa isang bagay - hindi sila bumubuo ng mga buto. Ang kanilang mga ugat ay epektibong sumisipsip ng tubig at mga mineral na asing-gamot, ang vascular system ng mga putot ay mapagkakatiwalaang dinala ang mga sangkap na kinakailangan para sa buhay sa lahat ng mga organo, at ang mga dahon ay aktibong nag-synthesize ng mga organikong sangkap. Kahit na ang mga spores ay bumuti at nakakuha ng isang malakas na shell ng selulusa. Hindi natatakot sa pagkatuyo, dinadala sila ng hangin sa mga malalayong distansya at hindi maaaring tumubo kaagad, ngunit pagkatapos ng isang tiyak na panahon ng dormancy (ang tinatawag na dormant spores). Gayunpaman, kahit na ang pinakaperpektong spore ay isang unicellular formation; hindi tulad ng mga buto, mabilis itong natutuyo at hindi naglalaman ng suplay ng mga sustansya, at samakatuwid ay hindi makapaghintay ng matagal para sa mga kanais-nais na kondisyon para sa pag-unlad. Ngunit ang pagbuo ng mga resting spores ay isang mahalagang milestone sa paraan upang magtanim ng mga halaman. Sa loob ng maraming milyun-milyong taon, ang klima sa ating planeta ay nanatiling mainit at mahalumigmig, ngunit ang ebolusyon sa matabang kagubatan ng mga bog ng karbon ay hindi huminto. Ang mga halamang may spore-bearing na puno ay unang nakabuo ng mga primitive na anyo ng mga tunay na buto. Lumitaw ang mga seed ferns (mga sikat na kinatawan ng genus Lepidodendron - sa Griyego ang pangalang ito ay nangangahulugang "scaly tree") at cordaites na may solid woody trunks. Bagama't kakaunti ang mga fossil ng mga organismong ito na nabuhay daan-daang milyong taon na ang nakalilipas, ang mga parang punong buto na pako ay kilala na umiral na bago pa ang Carboniferous. Noong tagsibol ng 1869, ang Skohary Creek sa Catskills, New York, ay bumaha nang husto. Inanod ng baha ang mga tulay, tinumba ang mga puno, at tinangay nang husto ang baybayin malapit sa nayon ng Gilboa. Ang pangyayaring ito ay matagal nang nakalimutan kung ang bumabagsak na tubig ay hindi nagsiwalat sa mga nagmamasid ng isang kahanga-hangang koleksyon ng mga kakaibang tuod. Ang kanilang mga base ay lumawak nang malaki, tulad ng mga puno ng swamp, ang diameter ay umabot sa 1.2 m, at ang kanilang edad ay 300 milyong taon. taon. Ang mga detalye ng istraktura ng bark ay mahusay na napanatili, ang mga fragment ng mga sanga at dahon ay nakakalat sa malapit. Naturally, ang lahat ng ito, kabilang ang silt mula sa kung saan ang mga tuod ay tumaas, ay petrified. Napetsahan sila ng mga geologist sa Upper Devonian, ang pre-Carboniferous period, at natukoy na tumutugma sila sa tree ferns. Sa susunod na limampung taon, tanging mga paleobotanist lamang ang naalala ang nahanap, at pagkatapos ay ang nayon ng Gilboa ay nagpakita ng isa pang sorpresa. Kasama ang mga fossilized trunks ng mga sinaunang pako, sa pagkakataong ito ay natuklasan ang kanilang mga sanga na may tunay na mga buto. Ngayon ang mga patay na punong ito ay nabibilang sa genus Eospermatopteris, na isinasalin bilang "dawn seed fern". ("liwayway", dahil pinag-uusapan natin ang tungkol sa pinakamaagang mga buto ng halaman sa Earth). Ang maalamat na panahon ng Carboniferous ay natapos nang ang mga geological na proseso ay nagpakumplikado sa lunas ng planeta, na nagdurog sa ibabaw nito sa mga tiklop at naghiwa-hiwalay nito sa mga hanay ng bundok. Ang mga latian sa mababang lupa ay inilibing sa ilalim ng isang makapal na patong ng mga sedimentary na bato na nahugasan sa mga dalisdis. Ang mga kontinente ay nagbago ng kanilang hugis, itinutulak ang dagat at lumihis mula sa dati nitong agos, ang mga takip ng yelo ay nagsimulang tumubo sa mga lugar, at ang pulang buhangin ay natakpan ang malawak na kalawakan ng lupa. Ang mga higanteng ferns, club mosses at horsetails ay namatay: ang kanilang mga spores ay hindi inangkop sa mas malupit na klima, at ang pagtatangkang lumipat sa pagpaparami sa pamamagitan ng mga buto ay naging masyadong mahina at hindi tiyak.
Ang unang tunay na binhing halaman. Ang mga kagubatan ng karbon ay nawala at natatakpan ng mga bagong layer ng buhangin at luad, ngunit ang ilang mga puno ay nakaligtas sa pamamagitan ng pagbuo ng mga buto na may pakpak na may isang malakas na shell. Ang gayong mga buto ay maaaring kumalat nang mas mabilis, mas mahaba, at samakatuwid ay sa mas mahabang distansya. Ang lahat ng ito ay nagpapataas ng kanilang mga pagkakataon na makahanap ng mga kanais-nais na kondisyon para sa pagtubo o paghihintay sa kanilang pagdating. Ang mga buto ay itinakda upang baguhin ang buhay sa Earth sa simula panahon ng mesozoic. Sa oras na ito, dalawang uri ng mga puno ang nakatakas sa malungkot na kapalaran ng iba pang carboniferous na mga halaman - at mga puno ng ginkgo. Ang mga pangkat na ito ay nagsimulang mag-co-populate sa mga kontinente ng Mesozoic. Hindi nakatagpo ng kumpetisyon, kumalat sila mula Greenland hanggang Antarctica, na ginagawang halos homogenous ang vegetation cover ng ating planeta. Ang kanilang mga buto na may pakpak ay naglakbay sa mga lambak ng bundok, lumipad sa ibabaw ng walang buhay na mga bato, umusbong sa mabuhangin na mga patch sa pagitan ng mga bato at sa pagitan ng alluvial na graba. Marahil, sila ay natulungan sa pagbuo ng mga bagong lugar sa pamamagitan ng maliliit at pako na nakaligtas sa pagbabago ng klima sa planeta sa ilalim ng mga bangin, sa lilim ng mga bangin at sa kahabaan ng baybayin ng mga lawa. Pinataba nila ang lupa gamit ang kanilang mga organikong labi, inihahanda ang matabang layer nito para sa pag-areglo ng mas malalaking species. Ang mga bulubundukin at malawak na kapatagan ay nanatiling hubad. Dalawang uri ng "pioneer" na puno na may pakpak na buto, na kumakalat sa buong planeta, ay itinali sa mga basang lugar, dahil ang kanilang mga itlog ay pinataba ng flagellated, aktibong lumalangoy na spermatozoa, tulad ng mga lumot at pako. marami halamang spore bumubuo ng mga spores ng iba't ibang laki - malalaking megaspores, na nagbibigay ng mga babaeng gametes, at maliliit na microspores, sa panahon ng paghahati kung saan lumitaw ang mga mobile. Upang lagyan ng pataba ang itlog, kailangan nilang lumangoy hanggang dito sa tubig - habang ang isang patak ng ulan at hamog ay sapat na. Sa cycads at ginkgoes, ang mga megaspores ay hindi nakakalat ng halaman ng magulang, ngunit nananatili dito, nagiging mga buto, ngunit sila ay mobile, kaya ang dampness ay kinakailangan para sa pagpapabunga. Panlabas na istraktura ng mga halamang ito, lalo na ang kanilang mga dahon, ay naglalapit din sa kanila sa mala-fern na mga ninuno. Ang pagpapanatili ng sinaunang paraan ng pagpapabunga sa pamamagitan ng spermatozoa na lumulutang sa tubig ay humantong sa katotohanan na, sa kabila ng medyo matitigas na mga buto, ang matagal na tagtuyot ay nanatiling isang hindi malulutas na problema para sa mga halaman na ito, at ang pagsakop sa lupa ay nasuspinde. Ang kinabukasan ng terrestrial vegetation ay ibinigay ng mga puno ng ibang uri, na tumubo sa mga cycad at ginkgo, ngunit nawala ang flagella. Ito ang mga araucaria (genus Araucaria), mga inapo ng Carboniferous cordaites, na nakaligtas hanggang ngayon. Sa panahon ng mga cycad, ang araucaria ay nagsimulang bumuo ng malaking halaga ng mga microscopic na butil ng pollen, na tumutugma sa microspores, ngunit tuyo at siksik. Dinala sila ng hangin sa mga megaspores, mas tiyak, sa mga ovule na nabuo mula sa kanila na may mga itlog, at tumubo sa mga pollen tubes na naghahatid ng hindi kumikilos na tamud sa mga babaeng gametes. Kaya, lumitaw ang pollen sa mundo. Ang pangangailangan para sa tubig para sa pagpapabunga ay nawala, at ang mga halaman ay tumaas sa isang bagong yugto ng ebolusyon. Ang pagbuo ng pollen ay humantong sa isang napakalaking pagtaas sa bilang ng mga buto na umuunlad sa bawat indibidwal na puno, at dahil dito sa mabilis na pagkalat ng mga halaman na ito. Ang sinaunang araucaria ay mayroon ding paraan ng pag-areglo, na napanatili sa mga modernong conifer, sa tulong ng mga matitigas na buto na may pakpak, na madaling dinala ng hangin. Kaya, ang una, at sa paglipas ng panahon, ang mga kilalang species ng pamilya ng pine ay lumitaw. Ang Pine ay gumagawa ng dalawang uri ng cones. Ang haba ng mga lalaki ay humigit-kumulang. Ang 2.5 cm at 6 na mm ang lapad ay pinagsama-sama sa mga dulo ng pinakamataas na sanga, kadalasan sa mga bungkos ng sampu o higit pa, upang ang isang malaking puno ay maaaring magkaroon ng ilang libo sa kanila. Sila ay nagkakalat ng pollen, na nagpapaligo sa lahat ng bagay sa paligid ng isang dilaw na pulbos. Ang mga babaeng cone ay mas malaki at lumalaki sa puno sa ibaba ng mga lalaki. Ang bawat isa sa kanilang mga kaliskis ay kahawig ng isang scoop sa hugis - malawak sa labas at patulis patungo sa base, kung saan ito ay nakakabit sa makahoy na axis ng kono. Sa itaas na bahagi ng sukat, mas malapit sa axis na ito, dalawang megaspores ay bukas na matatagpuan, naghihintay para sa polinasyon at pagpapabunga. Ang mga butil ng pollen na dala ng hangin ay lumilipad sa loob ng mga babaeng cone, gumulong pababa sa mga kaliskis hanggang sa mga ovule at nakipag-ugnayan sa kanila, na kinakailangan para sa pagpapabunga. Ang mga cycad at ginkgo ay hindi maaaring makipagkumpitensya sa mas progresibong mga conifer, na, epektibong nagpapakalat ng pollen at may pakpak na mga buto, hindi lamang nagtulak sa kanila palabas, ngunit pinagkadalubhasaan din ang mga bago, dati nang hindi naa-access na mga sulok ng lupain. Ang mga taxodies ay naging unang coniferous dominant (ngayon kasama na nila, sa partikular, sequoias at marsh). Ang pagkakaroon ng pagkalat sa buong mundo, ang magagandang punong ito ay sumasakop sa lahat ng bahagi ng mundo na may pare-parehong mga halaman sa huling pagkakataon: ang kanilang mga labi ay matatagpuan sa Europa, Hilagang Amerika, Siberia, China, Greenland, Alaska at Japan.
halaman at ang kanilang mga buto., at Ginkgo ay nabibilang sa tinatawag na. mga gymnosperm. Nangangahulugan ito na ang kanilang mga ovule ay bukas na matatagpuan sa mga kaliskis ng buto. Ang mga halaman ay bumubuo sa departamento ng mga angiosperms: ang kanilang mga ovule at ang mga buto na umuunlad mula sa kanila ay nakatago mula sa panlabas na kapaligiran sa pinalawak na base ng pistil, na tinatawag na ovary. Bilang resulta, ang butil ng pollen ay hindi direktang maabot ang ovule. Para sa pagsasanib ng mga gametes at pag-unlad ng buto, kinakailangan ang isang ganap na bagong istraktura ng halaman - isang bulaklak. Ang bahaging lalaki nito ay kinakatawan ng mga stamen, ang bahagi ng babae ay mga pistil. Maaari silang maging sa parehong bulaklak o sa iba't ibang mga bulaklak, kahit na sa iba't ibang mga halaman, na sa huling kaso ay tinatawag na dioecious. Kabilang sa mga dioecious species, halimbawa, ash, holly, poplar, willow, petsa. Para maganap ang pagpapabunga, ang butil ng pollen ay dapat dumapo sa tuktok ng pistil—ang malagkit, minsan pinnate na stigma—at sumunod dito. Ang stigma ay naglalabas ng mga kemikal, sa ilalim ng impluwensya kung saan ang butil ng pollen ay tumutubo: ang buhay na protoplasm, na lumalabas mula sa ilalim ng matigas na shell nito, ay bumubuo ng isang mahabang pollen tube na tumagos sa stigma, kumakalat nang mas malalim sa pistil kasama ang pahabang bahagi nito (column) at sa huli. umabot sa obaryo na may mga ovule. Sa ilalim ng impluwensya ng mga chemical attractant, ang nucleus ng male gamete ay gumagalaw kasama ang pollen tube hanggang sa ovule, tumagos dito sa pamamagitan ng isang maliit na butas (micropyle) at sumasama sa nucleus ng itlog. Ito ay kung paano nangyayari ang pagpapabunga. Pagkatapos nito, ang buto ay nagsisimulang umunlad - sa isang mahalumigmig na kapaligiran, sagana na ibinibigay sa mga sustansya, na protektado ng mga dingding ng obaryo mula sa mga panlabas na impluwensya. Ang magkatulad na pagbabagong ebolusyon ay kilala rin sa mundo ng hayop: ang panlabas na pagpapabunga, na karaniwan, halimbawa, para sa mga isda, ay pinalitan ng panloob na pagpapabunga sa lupa, at ang embryo ng mga mammal ay nabuo hindi sa mga itlog na inilatag sa panlabas na kapaligiran, bilang, para sa halimbawa, sa mga karaniwang reptilya, ngunit sa loob ng matris. Ang paghihiwalay ng umuunlad na binhi mula sa mga panlabas na impluwensya ay nagpapahintulot sa mga namumulaklak na halaman na matapang na "mag-eksperimento" sa hugis at istraktura nito, at ito naman, ay humantong sa isang mala-avalanche na hitsura ng mga bagong anyo. halaman sa lupa, ang pagkakaiba-iba nito ay nagsimulang lumago sa hindi pa nagagawang bilis sa mga nakaraang panahon. Ang kaibahan sa gymnosperms ay halata. Ang kanilang "hubad" na mga buto na nakahiga sa ibabaw ng mga kaliskis, anuman ang uri ng halaman, ay humigit-kumulang pareho: hugis ng patak ng luha, natatakpan ng matigas na balat, kung saan ang isang patag na pakpak ay minsan nakakabit, na nabuo ng mga selulang nakapalibot sa buto. Hindi nakakagulat na sa maraming milyong taon ang anyo ng gymnosperms ay nanatiling napakakonserbatibo: ang mga pine, spruces, firs, yews, ay halos magkapareho sa bawat isa. Totoo, sa junipers, yew at ginkgo seeds ay maaaring malito sa mga berry, ngunit hindi nito binabago ang pangkalahatang larawan - ang matinding pagkakapareho ng pangkalahatang plano ng istraktura ng gymnosperms, ang laki, uri at kulay ng kanilang mga buto kumpara sa malaking kayamanan ng mga anyo ng pamumulaklak. Sa kabila ng kakulangan ng impormasyon tungkol sa mga unang yugto ng ebolusyon ng angiosperms, pinaniniwalaan na lumitaw ang mga ito sa pagtatapos ng panahon ng Mesozoic, na natapos mga 65 milyong taon na ang nakalilipas, at sa simula. panahon ng cenozoic nasakop na ang mundo. Ang pinakalumang kilalang genus ng pamumulaklak ay Claytonia. Natagpuan ito sa Greenland at Sardinia, ibig sabihin, malamang na kahit na 155 milyong taon na ang nakalilipas ito ay kasing laganap. Ang mga dahon ng Claytonia ay palmately complex, tulad ng mga kasalukuyang horse chestnut at lupins, at ang mga prutas ay parang berry na may diameter na 0.5 cm sa dulo ng manipis na peduncle. Marahil ang mga halamang ito ay kayumanggi o berde ang kulay. Ang mga maliliwanag na kulay ng mga bulaklak at prutas ng angiosperm ay lumitaw sa ibang pagkakataon - kaayon ng ebolusyon ng mga insekto at iba pang mga hayop, na idinisenyo upang maakit. Ang berry ng Claytonia ay apat na binhi; dito makikita mo ang isang bagay na kahawig ng labi ng isang mantsa. Bilang karagdagan sa napakabihirang mga labi ng fossil, ang hindi pangkaraniwang modernong mga halaman, na naka-grupo sa order na Gnetales, ay nagbibigay ng ilang pananaw sa mga unang namumulaklak na halaman. Ang isa sa kanilang mga kinatawan ay ang conifer (genus Ephedra), na natagpuan, sa partikular, sa mga disyerto ng timog-kanluran ng Estados Unidos; sa panlabas, ito ay parang ilang walang dahon na mga tungkod na umaabot mula sa isang makapal na tangkay. Ang isa pang genus - velvichia (Welwitschia) ay lumalaki sa disyerto sa timog-kanlurang baybayin ng Africa, at ang pangatlo - gnetum (Gnetum) - isang mababang palumpong ng tropiko ng India at Malay. Ang tatlong genera na ito ay maaaring ituring na "mga nabubuhay na fossil", na nagpapakita ng mga posibleng landas ng pagbabago gymnosperms sa angiosperms. Ang mga cone ng conifer ay panlabas na kahawig ng mga bulaklak: ang kanilang mga kaliskis ay nahahati sa dalawang bahagi, na kahawig ng mga petals. Ang Velvichia ay mayroon lamang dalawang malapad na laso na tulad ng mga dahon hanggang sa 3 m ang haba, ganap na naiiba sa mga karayom ng conifer. Ang mga buto ng gnetum ay binibigyan ng karagdagang shell, na ginagawa itong parang angiosperm drupes. Ito ay kilala na ang angiosperms ay naiiba mula sa gymnosperms sa istraktura ng kahoy. Sa mga mapang-api, pinagsasama nito ang mga katangian ng magkabilang grupo.
Pagpapakalat ng binhi. Ang posibilidad at pagkakaiba-iba ng mundo ng halaman ay nakasalalay sa kakayahan ng mga species na kumalat. Ang magulang na halaman ay nakaugat sa isang lugar sa buong buhay nito, samakatuwid, ang mga supling nito ay dapat makahanap ng isa pa. Ang gawaing ito ng pagbuo ng bagong espasyo ay ipinagkatiwala sa mga buto. Una, ang pollen ay dapat mapunta sa pistil ng isang bulaklak ng parehong species, i.e. dapat maganap ang polinasyon. Pangalawa, ang pollen tube ay dapat umabot sa ovule, kung saan ang nuclei ng male at female gametes ay magsasama. Sa wakas, ang mature na binhi ay kailangang umalis sa magulang na halaman. Ang posibilidad na ang isang buto ay tumubo at matagumpay na mag-ugat sa isang bagong lugar ay isang maliit na bahagi ng isang porsyento, kaya ang mga halaman ay napipilitang umasa sa batas ng malaking bilang at ikalat ang pinakamaraming buto hangga't maaari. Ang huling parameter ay karaniwang inversely proportional sa kanilang mga pagkakataong mabuhay. Ihambing natin, halimbawa, ang isang niyog at mga orchid. Ang niyog ang may pinakamalaki flora mga buto. Nagagawa nilang lumangoy nang walang hanggan sa mga karagatan hanggang sa itapon sila ng mga alon sa malambot na buhangin sa baybayin, kung saan ang kumpetisyon ng mga punla sa iba pang mga halaman ay magiging mas mahina kaysa sa kagubatan nang mas madalas. Bilang isang resulta, ang mga pagkakataon na manirahan para sa bawat isa sa kanila ay medyo mataas, at ang isang mature na walang panganib sa mga species ay kadalasang nagdadala lamang ng ilang dosenang mga buto bawat taon. Ang mga orchid naman ay may pinakamaliit na buto sa mundo; sa mga tropikal na kagubatan, dinadala sila ng mahinang agos ng hangin sa mga matataas na korona at tumutubo sa basa-basa na mga bitak sa balat sa mga sanga ng puno. Ang sitwasyon ay kumplikado sa pamamagitan ng katotohanan na sa mga sanga na ito kailangan nilang makahanap ng isang espesyal na uri ng fungus, kung wala ang pagtubo ay imposible: ang mga maliliit na buto ng orchid ay hindi naglalaman ng mga reserbang nutrisyon at sa mga unang yugto ng pag-unlad ng punla ay natatanggap nila ang mga ito mula sa fungus. Hindi kataka-taka na sa isang prutas ng isang maliit na orkidyas mayroong ilang libong mga butong ito. angiosperms ay hindi limitado sa pagbuo ng iba't ibang mga buto bilang resulta ng pagpapabunga: ang mga ovary, at kung minsan ang iba pang mga bahagi ng mga bulaklak, ay bubuo sa mga natatanging istruktura na naglalaman ng mga buto - mga prutas. Ang obaryo ay maaaring maging isang green bean na nagpoprotekta sa mga buto hanggang sa sila ay mahinog, maging isang malakas na niyog na may kakayahang gumawa ng mahabang paglalakbay sa dagat, maging isang makatas na mansanas na kakainin ng isang hayop sa isang liblib na lugar, gamit ang pulp, ngunit hindi ang mga buto. Ang mga berry at drupes ay isang paboritong delicacy ng mga ibon: ang mga buto ng mga prutas na ito ay hindi natutunaw sa kanilang mga bituka at nahuhulog sa lupa kasama ng dumi, kung minsan ay maraming kilometro mula sa halaman ng magulang. Ang mga prutas ay may pakpak at mahimulmol, at ang anyo ng mga appendage na nagpapataas ng pagkasumpungin ay mas magkakaibang kaysa sa mga buto ng pine. Ang pakpak ng prutas na abo ay kahawig ng isang sagwan, sa elm ay tila isang labi ng isang sumbrero, sa maple ang magkapares na mga prutas - diptera - ay kahawig ng mga ibon na lumulutang, sa ailanthus ang mga pakpak ng prutas ay pinaikot sa isang anggulo sa bawat isa. iba pa, na bumubuo, bilang ito ay, isang propeller. Ang mga adaptasyon na ito ay nagpapahintulot sa mga namumulaklak na halaman na gumamit ng mga panlabas na salik nang napakabisa para sa pagpapakalat ng mga buto. Gayunpaman, ang ilang mga species ay hindi umaasa sa tulong sa labas. Kaya, ang mga bunga ng touchy ay isang uri ng tirador. Gumagamit ang mga geranium ng katulad na mekanismo. Sa loob ng kanilang mahabang prutas, isang baras ang dumaan, kung saan apat, pansamantala, ang mga tuwid at pinagsama-samang mga flap ay nakakabit - mahigpit silang humahawak mula sa itaas, mahina mula sa ibaba. Kapag hinog na, ang mga ibabang dulo ng mga balbula ay lumalabas sa base, umiikot nang husto patungo sa tuktok ng baras at ikalat ang mga buto. Sa ceanothus shrub na kilalang-kilala sa Amerika, ang obaryo ay nagiging isang berry, na katulad ng istraktura sa isang bomba ng oras. Ang presyon ng katas sa loob ay napakataas na pagkatapos ng pagkahinog, ang isang sinag ng sikat ng araw ay sapat na mainit upang ang mga buto nito ay nakakalat sa lahat ng direksyon tulad ng mga buhay na shrapnel. Mga kahon ng ordinaryong violet, na natuyo, sumabog at nagkalat ng mga buto sa kanilang paligid. Ang mga prutas ng Hamamelis ay kumikilos tulad ng isang howitzer: upang ang mga buto ay mahulog nang mas malayo, sila ay kukunan ang mga ito sa isang mataas na anggulo sa abot-tanaw. Sa knotweed virgin, sa lugar kung saan nakakabit ang mga buto sa halaman, nabuo ang isang tulad ng spring na istraktura na nagtatapon ng mga mature na buto. Sa oxalis, ang mga shell ng prutas ay unang namamaga, at pagkatapos ay pumutok at lumiliit nang husto na ang mga buto ay lumilipad sa mga bitak. Ang Arceutobium ay maliit, dahil sa haydroliko na presyon sa loob ng mga berry, itinutulak nito ang mga buto palabas ng mga ito tulad ng mga miniature na torpedo.

PAHAGI NG MGA BINHI NA MAY TUBIG>">
> Pag-unlad ng buto mula sa ovule. Istraktura ng buto. Mga uri ng binhi. Biyolohikal na papel at pang-ekonomiyang paggamit ng mga buto
plastid chloroplast mitochondrion phylogeny
Binhi - isang espesyal na multicellular na istraktura ng isang kumplikadong istraktura, na nagsisilbi para sa pagpaparami at pag-aayos ng mga halaman ng buto, kadalasang umuunlad pagkatapos ng pagpapabunga mula sa ovule (isang binagong babaeng sporangium) at naglalaman ng isang embryo.
Bagama't ang binhi ay madalas na inilarawan (kabilang ang mga mapagkukunang may awtoridad) bilang isang "organ pagpaparami ng binhi halaman "(mas madalas -" ang organ ng sekswal na pagpaparami ng mga halaman "), ang buto ay hindi isang organ sa karaniwang kahulugan ng terminong ito, dahil pinagsasama nito ang mga istruktura ng dalawa (sa gymnosperms - tatlo) magkakaibang henerasyon ikot ng buhay. Ang mga organo ng sekswal na pagpaparami (genital organ, gametangia) sa gymnosperms ay kinakatawan ng archegonia, habang sa mga namumulaklak na halaman sila ay nabawasan. Ang higit na makatwiran ay ang kahulugan ng isang buto bilang isang "pangunahing halaman" (ito ay ibinigay ng maraming mga aklat-aralin sa paaralan ng botany); binibigyang-diin ng kahulugang ito na ang isang bagong henerasyon (sporophyte) ng halaman ay bubuo mula sa binhi. Kasabay nito, ang natitirang bahagi ng buto, maliban sa embryo, ay maaaring ituring na mga karagdagang istruktura (organ) na nagsisiguro sa pag-unlad ng embryo.
Istraktura ng buto
Ang buto ay bubuo sa ibabaw ng seed scale. Ito ay isang multicellular na istraktura na pinagsasama ang storage tissue - endosperm, embryo at isang espesyal na proteksiyon na takip (seed peel). Bago ang pagpapabunga, ang gitnang bahagi ng ovule ay naglalaman ng nucellus, na unti-unting pinalitan ng endosperm. Ang endosperm ay haploid at nabuo mula sa mga tisyu ng babaeng gametophyte.
Sa cycads at ginkgoes, ang panlabas na layer ng seed coat (sarcotesta) ay malambot at mataba, ang gitnang layer (sclerotesta) ay matigas, at ang panloob na layer (endotesta) ay may lamad sa oras na ang buto ay hinog. Ang mga buto ay dispersed ng iba't ibang mga hayop na kumakain ng sarcotesta nang hindi nakakasira sa sclerotesta.
Sa yew at podocarpus, ang mga buto ay napapalibutan ng isang mataba na aryllus, isang lubos na binagong sukat ng babaeng kono. Ang makatas at maliwanag na kulay na arillus ay umaakit sa mga ibon na nagkakalat ng mga buto ng mga conifer na ito. Ang Arillus ng maraming species ng podocarpus ay nakakain din para sa mga tao.
Ang istraktura ng mga buto ng angiosperms. Testa.
Sa labas, ang binhi ay natatakpan ng isang seed coat, na nagpoprotekta sa mga panloob na bahagi ng buto mula sa pagkatuyo at mekanikal na pinsala. Ang seed coat ay bubuo mula sa integument ng ovule.
Ang endosperm ay ang tissue na nakapaloob sa loob ng buto, kadalasang nakapalibot sa embryo at nagbibigay ito ng mga sustansya sa panahon ng pag-unlad. Sa gymnosperms, ang endosperm ay ang tissue ng babaeng gametophyte. Kadalasan sa mga unang yugto ng pag-unlad, mayroon itong syncytial na istraktura, mamaya mga pader ng cell. Ang mga endosperm cell ay haploid sa una ngunit maaaring maging polyploid. Sa mga namumulaklak na halaman, ang endosperm ay karaniwang nabuo sa panahon ng dobleng pagpapabunga bilang resulta ng pagsasanib ng central cell (central nucleus) ng embryo sac sa isa sa sperm. Sa maraming mga namumulaklak na selula, ang endosperm ay triploid. Sa isang water lily, ang endosperm ay nabuo sa pamamagitan ng pagsasanib ng tamud na may isang haploid cell ng embryo sac, upang ang nuclei nito ay diploid. Sa maraming flower nuclei, ang endosperm ay may chromosome set na higit sa 3n (hanggang 15n).
Ang perisperm ay katulad sa pag-andar sa endosperm, ngunit may isang diploid na hanay ng mga chromosome, naglalaman ng isang maliit na halaga ng mga protina, pangunahin ang starch, at kung minsan ay mga taba. Maaari nitong gampanan ang papel ng pangunahing imbakan na tisyu kapwa nang nakapag-iisa at kasama ng endosperm.
Sa ilalim ng alisan ng balat ay isang embryo - isang maliit na halaman sa hinaharap. Ang embryo sa maraming namumulaklak na halaman ay binubuo ng germinal root, germinal stalk, germinal bud, at cotyledon. Sa ibang mga grupo (halimbawa, sa napakaraming orchid), ang embryo ay walang magkakaibang mga organo bago ang pagtubo ng buto.
Mga tampok ng istraktura ng mga buto ng monocots at dicots
Laki ng buto
Ang mga buto ng orchid at broomrape ay may pinakamababang sukat; ang kanilang masa ay 0.001-0.003 mg. Ang mga buto ng Seychelles palm Lodoicea maldivica ay umabot sa pinakamataas na sukat (timbang na mga 20 kg).
Kemikal na komposisyon ng mga buto
Ang pangunahing tampok ng kemikal na komposisyon ng mga mature na buto ay isang napakababang nilalaman ng tubig, karaniwang 10-15% lamang (ayon sa iba't ibang mga mapagkukunan, mula 5 hanggang 20%).
Ang kemikal na komposisyon ng mga buto ay lubos na nakasalalay sa mga kondisyon ng pagkahinog at sa iba't ibang halaman. Ang mga buto ng ilang mga halaman ay naglalaman ng higit pang mga protina (beans, soybeans), ang iba pa - mas maraming carbohydrates (trigo, rye), iba pa - mas maraming taba (sunflower, nuts). Ang mga enzyme (maltase, lipase, phosphatase, proteolytic) ay kinakailangan para sa pag-convert ng mga reserbang nutrients sa isang form na natutunaw para sa embryo.
Ang pagtubo ng binhi ay ang kanilang kakayahang magbigay ng normal na mga punla (sa laboratoryo) o mga punla (sa bukid) sa isang tiyak na tagal ng panahon. Ang pagsibol ay lubos na nakasalalay sa mga kondisyon ng pagtubo at mga kondisyon ng imbakan ng mga buto. Ang pagtubo ay karaniwang ipinapahayag bilang isang porsyento (ito ang porsyento ng mga buto na tumubo mula sa kabuuang bilang ng mga buto).
Sa pangmatagalang imbakan ng mga buto, bumababa ang kanilang pagtubo sa paglipas ng panahon. Ang mga buto ng ilang mga halaman ay nawawalan ng kakayahang mabuhay pagkatapos ng 2-3 linggo (halimbawa, ang mga buto ng karamihan sa mga species ng willow ay ganap na nawawalan ng kakayahang mabuhay sa temperatura na 18-20°C sa loob ng isang buwan). Karamihan sa pagtubo ng binhi mga nilinang na halaman kapansin-pansing nabawasan pagkatapos ng 2-3 taon. Ang mga buto ng lotus sa pit ay nananatiling mabubuhay nang hindi bababa sa 250 taon (ayon sa ilang mga mapagkukunan, higit sa isang libong taon). Ang mga buto ng Arctic lupine na napanatili sa permafrost ay pinamamahalaang tumubo pagkatapos ng 10-12 libong taon.
kondisyon ng pagtubo ng binhi
Ang mga buto ng halaman ay tumubo sa isang positibong temperatura. Ang temperatura kung saan nagsisimula ang pagtubo ay malawak na nag-iiba-iba sa mga halaman ng iba't ibang pangkat ng taxonomic at heyograpikong rehiyon. Sa karaniwan, ang mga buto ng mga halaman ng polar at temperate latitude ay tumutubo sa mas mababang temperatura kaysa sa mga buto ng subtropikal at tropikal na species. Ang pinakamainam na temperatura ng pagtubo ay iba rin, kung saan ang pinakamalaking pagtubo at pinakamataas na pagtubo ay sinusunod.
Ang mga buto ng ilang mga halaman ay nakatiis sa mga panahon ng panandaliang pagkakalantad sa mataas na temperatura sa panahon ng mga sunog sa kagubatan, pagkatapos ay nilikha ang mga ito kanais-nais na mga kondisyon para sa pagtubo ng mga nabubuhay na binhi. Bilang karagdagan, ang apoy ay nakakatulong sa pagbubukas ng mga bunga ng ilang uri ng halaman na lumalaban sa apoy. Kaya, pagkatapos lamang ng mga apoy ay ang "huli" na mga cone ng lodgepole pine, ang mga cone ng sequoiadendron, atbp., Ang mga bunga ng ilang mga species ng genus Banksia, ay binuksan.
Stratification - pinapanatili ang mga buto sa mababang positibong temperatura.
Scarification - pinsala sa pamamagitan ng mekanikal o kemikal na pagkilos ng seed coat, kinakailangan para sa kanilang pagtubo. Ito ay karaniwang kinakailangan para sa mga buto na may makapal at matibay na seed coat (maraming legume) o endocarp (halimbawa, raspberry, bird cherry).
Sa likas na katangian, ang pagkakalantad sa bakterya at humic acid sa lupa, pati na rin ang pagpasa sa gastrointestinal tract ng iba't ibang mga hayop, ay maaaring magsilbi bilang isang scarifying agent.
Ipinapalagay na ang mga buto ng ilang mga halaman (halimbawa, calvaria Sideroxylon grandiflorum) ay hindi maaaring tumubo sa kalikasan nang hindi dumadaan sa mga bituka ng mga ibon. Kaya, ang mga buto ng calvaria ay maaaring tumubo lamang pagkatapos nilang dumaan sa mga bituka ng mga domestic turkey o tratuhin ng polishing paste.
Ang ilang mga buto ay nangangailangan ng parehong scarification at stratification sa parehong oras. At kung minsan (hawthorn) karamihan sa mga buto ay tumutubo pagkatapos ng scarification at double stratification, iyon ay, pagkatapos ng dalawang taglamig na natutulog na panahon.
Ang papel ng mga buto sa kalikasan at buhay ng tao
Maraming mga organismo (mula sa fungi at bakterya hanggang sa mga ibon at mammal) ay kumakain nang husto, at kung minsan ay eksklusibo, sa mga buto. Binubuo ng mga buto ang batayan ng pagkain ng mga hayop tulad ng ilang mga insekto at kanilang larvae (halimbawa, reaper ants), granivorous na ibon, rodents (chipmunks, squirrels, hamster, atbp.).
Mula sa simula ng agrikultura sa karamihan ng mga rehiyon ng mundo, ang batayan ng pagkain ng tao ay mga buto din, pangunahin ng mga nilinang cereal (trigo, bigas, mais, atbp.). Ang pangunahing nutrient na natatanggap ng sangkatauhan pinakamalaking bilang calories, - almirol na nakapaloob sa mga buto ng cereal. Ang isang mahalagang pinagmumulan ng mga protina para sa sangkatauhan ay ang mga buto din ng mga leguminous na halaman - soybeans, beans, atbp. Ang mga buto ay ang pangunahing pinagkukunan ng mga langis ng gulay, na nakuha mula sa mga buto ng mirasol, rapeseed, mais, flax, koton at marami pang ibang mga oilseed.
Mula sa sandali ng pagsisimula hanggang sa ganap na pagkahinog, kapag ito ay naging may kakayahang gumawa ng isang normal na usbong, ang isang binhi ay dumaan sa isang serye ng mga kumplikadong pagbabago mula sa isang estado patungo sa isa pa, mas perpekto, iyon ay, kung ano ang nangyayari ay kung ano ang tinukoy ng konsepto. ng "pag-unlad ng binhi".
Ang buong masalimuot na proseso ay maaaring hatiin sa ilang mga panahon at mga yugto na nagpapakilala sa mga indibidwal na yugto sa buhay ng mga buto.
Ang bawat yugto ay may ganap na tiyak na estado ng binhi, at samakatuwid ang diagnosis ng yugto ay dapat na makilala sa pamamagitan ng sukdulang kalinawan at pagiging simple. Gayunpaman, ngayon ay mayroon lamang mga nakakalat na paglalarawan ng mga indibidwal na yugto, kadalasan sa anumang batayan.
Ang pag-uuri ng mga panahon at yugto ng pag-unlad ng binhi ay lalong mahalaga. Upang bumuo ng isang pag-uuri ng ito o ang hindi pangkaraniwang bagay na iyon, kinakailangan na gawing pangkalahatan ang naipon na pang-eksperimentong materyal at ibuod ang mga resulta ng pananaliksik at magmungkahi ng isang paraan para sa karagdagang pag-unlad ng hindi pangkaraniwang bagay na ito. Naturally, ang ganitong pag-uuri ay mabubuo lamang sa pamamagitan ng sama-samang pagsisikap ng mga mananaliksik.
Ang batayan para sa pagbuo ng isang pag-uuri ng mga panahon at mga yugto ng pag-unlad ng binhi ay dapat na isang hanay ng mga tampok: morphological, morphogenetic at biochemical.
Ang mga yugto ay pinag-aralan sa pinakadetalye at ang mga pag-uuri ayon sa mga pananim na butil ay binuo. Ang pinakamahusay na mga klasipikasyon para sa mga pananim ng butil ay iminungkahi ni N. N. Kuleshov, para sa mga legume - ni V. A. Vishnevsky, para sa mga sunflower - ni V. K. Morozov.
Mga panahon ng pag-unlad ng binhi
Ang panahon ng pag-unlad ng binhi ay nailalarawan sa pamamagitan ng ilang makabuluhang pagbabago sa husay, pati na rin ang tagal nito.
Para sa mga pananim na butil, anim na katangian, malinaw na tinukoy na mga panahon ay maaaring makilala: pagbuo ng binhi(embryonic), pagbuo, pagpupuno, pagkahinog, post-harvest ripening, ganap na pagkahinog. Tulad ng makikita natin sa ibang pagkakataon, ang lahat ng mga panahong ito sa isang pangkalahatang anyo ay likas sa lahat ng iba pang mga kultura, bagaman, natural, ang bawat kultura ay magkakaroon ng mga tiyak na pagkakaiba sa likas na katangian ng panahon, sa mga yugto nito.
Hinati ni N. N. Kuleshov ang proseso ng pagbuo ng butil sa tatlong yugto (phase): pagbuo, pagpupuno At pagkahinog. Nakikita namin ang huling dalawang panahon sa interpretasyon ng N. N. Kuleshov, at hinati namin ang unang panahon sa dalawang magkaibang husay na mga panahon: pagbuo ng binhi at ang kanyang pagbuo. Bilang karagdagan, isinama namin sa isang proseso ng pagbuo ng binhi ang panahon post-harvest ripening at panahon ganap na pagkahinog.
Ang lahat ng mga panahong ito ay maaaring mailarawan bilang mga sumusunod (sa halimbawa ng trigo ng taglamig).
Panahon ng pagbuo ng binhi ay nagsisimula pagkatapos ng pagpapabunga (mula sa simula ng postgamous phase) at nagpapatuloy hanggang sa sandaling ang binhi, na nahiwalay sa inang halaman, ay maaaring umusbong. Ito ay nagpapahiwatig na ang binhi ay nabuo na at sa hinaharap sa isang panahon ng pagpapalakas nito, magsisimula ang pagbuo nito. Ang panahon ng embryonic na ito ay nagsisimula sa pagbuo ng zygote at nagtatapos sa pagbuo ng growth point ng embryo. Sa ganitong estado, ang embryo ay magagawa, sa ilalim ng pinakamainam na mga kondisyon, upang magbigay ng isang mahina, ngunit mabubuhay pa rin sprout.
Ang panahong ito ay tumatagal ng 7-9 araw para sa winter wheat, 7 araw para sa soft spring wheat, 10 araw para sa hard spring wheat, 10-15 araw para sa mais, atbp.
Panahon ng pagbuo nagpapatuloy hanggang sa maabot ang huling katangian ng haba ng butil ng iba't-ibang ito. Sa pagtatapos ng panahon, ang pagkakaiba-iba ng embryo ay karaniwang tapos na. Sa panahong ito, ang nilalaman ng butil ay nagiging gatas mula sa tubig (lumalabas ang mga butil ng starch sa endosperm tissue), at ang kulay ng shell ay nagbabago mula puti hanggang berde (naiipon ang chlorophyll). Ang moisture content ng butil ay 65-80%, at ang dry weight ng 1000 grains ay umabot sa 8-12 g. Ang panahong ito sa pag-unlad ng butil ay nailalarawan sa pamamagitan ng mataas na nilalaman ng tubig (lalo na ang libreng tubig) at isang mababang nilalaman ng tuyong bagay . Ang panahon ay tumatagal ng 5-8 araw.
Panahon ng pagpuno nagsisimula sa pagdeposito ng starch sa mga endosperm cells at nagpapatuloy hanggang sa tumigil ang pagdeposito ng starch. Ang panahon ay nailalarawan sa pamamagitan ng pagtaas sa lapad at kapal ng butil sa pinakamataas na sukat, ang kumpletong pagbuo ng endosperm tissue, na una ay may milky, pagkatapos ay pasty, at waxy consistency sa pagtatapos ng period. Ang bigat ng tubig sa butil ay nananatiling pare-pareho, ngunit ang moisture content ng butil ay nabawasan sa 38-40% (dahil sa patuloy na pagtaas ng dry matter). Ang panahong ito ay tumatagal ng isang average ng 20-25 araw, ngunit sa basa at malamig na panahon maaari itong tumagal ng hanggang 30 araw, at sa tuyo at mainit na panahon maaari itong bawasan sa 15-18 araw o mas kaunti.
panahon ng pagkahinog ng binhi nagsisimula sa pag-alis nito mula sa inang halaman, kapag huminto ang daloy ng mga plastik na sangkap, enzymes at maging ang tubig. Sa butil mayroong mga proseso ng polimerisasyon at pagpapatayo. Ang kahalumigmigan sa oras na ito ay bumababa sa 12-18%, at kung minsan hanggang 8%. Ang dami ng libreng tubig ay nabawasan nang husto, at sa pagtatapos ng panahon maaari itong mawala nang lubusan.
Ang nasabing paghahati sa mga panahon ay tama mula sa punto ng view ng mabibili na butil - ang huli ay tumatanda at itinuturing na angkop para sa teknikal na paggamit, iyon ay, ito ay nagiging isang hilaw na materyal para sa industriya.
Mula sa punto ng view ng seed grower, ang pagbuo ng mga buto ay hindi pa nakumpleto sa panahong ito. Tulad ng makikita natin sa ibang pagkakataon, nagsisimula ang isang bagong panahon ng husay, na nauugnay sa isang karagdagang pagbabago mga kemikal na sangkap at ang hitsura ng isang bago at pinakamahalagang pag-aari ng mga buto - ganap na normal na pagtubo. Kahit na ang morphological formation ng mga buto ay nagtatapos sa ikatlong panahon, ngunit ang physiological na proseso ay nagpapatuloy sa kasunod na panahon, samakatuwid, isinasaalang-alang namin na kinakailangan upang madagdagan ang proseso ng pagbuo ng binhi sa ikalimang panahon - ang panahon. post-harvest ripening.
SA panahon post-harvest ripening ang mga buto ay sumasailalim sa kumplikadong biochemical transformations ng iba't-ibang mga kemikal na compound, Bagama't mga tampok na morphological mananatiling pareho sa nakaraang yugto.
Sa panahong ito, ang synthesis ng mga high-molecular protein compound ay nagpapatuloy at nagtatapos, ang conversion ng mga libreng fatty acid sa taba, ang mga molekula ng mga compound ng carbohydrate ay lumalaki, ang mga proseso ng pagbabagong-anyo ng mga sangkap - mga inhibitor ng pagtubo sa iba pang mga anyo ay nagaganap, ang aktibidad. ng enzymes fades, hangin at tubig permeability ng seed coats ay tumataas.
Ang halumigmig ng buto ay nasa ekwilibriyo sa relatibong halumigmig ng hangin. Ang paghinga ng buto ay kumukupas. Sa simula ng panahon, ang mga buto ay hindi tumubo o ang kanilang pagtubo ay napakababa, sa dulo ito ay nagiging normal. Ang panahon ay tumatagal depende sa kultura at panlabas na mga kondisyon mula sa isang araw hanggang ilang buwan.
buong panahon ng pagkahinog ay nagsisimula sa simula ng buong pagtubo ng mga buto, iyon ay, ang mga buto ay handa na upang simulan ang isang bagong cycle sa buhay ng halaman. Mayroong mabagal na pagtanda ng mga colloid, na sinamahan ng mahinang paghinga. Sa ganitong estado, ang mga buto ay hanggang sa simula ng pagtubo o hanggang sa kumpletong kamatayan dahil sa pagtanda sa panahon ng pangmatagalang imbakan.
Sa ilang mga kaso, ang mga panahong ito ay nahahati sa mas maliliit na yugto ng pag-unlad ng binhi - mga yugto . Ang mga yugto ay nakikilala sa pamamagitan ng iba't ibang mga tampok, na pinakamalinaw na sumasalamin sa kanilang kakaiba. Sa isang kaso, ito ay maaaring isang espesyal na estado ng endosperm, sa isa pa, ang likas na katangian ng mga proseso ng physiological, atbp.
Ang panahon ng pagpuno ay nahahati sa mga sumusunod na yugto ng pag-unlad ayon sa estado ng endosperm: matubig, pre-dairy, pagawaan ng gatas, matamis. Sa panahon ng ripening, ang mga yugto ng pagkahinog ay nakikilala: waxy(madalas na makilala sa pagitan ng simula, puno at katapusan ng waxy ripeness), mahirap(minsan markahan ang simula ng solid phase ng pagkahinog).
Matubig na yugto- ang simula ng pagbuo ng mga endosperm cells. Ang butil ay puno ng tubig na likido. Ang shell ay puti o maputi-puti. Ang kahalumigmigan ng butil ay 75-80%, ang libreng kahalumigmigan ay 5-6 beses na higit pa kaysa sa nakagapos na kahalumigmigan, ang tuyong bagay ay 2-3% ng maximum na halaga. Ang tagal ng yugto ay nasa average na mga 6 na araw.
Pre-milk phase- ang likido, matubig na nilalaman ng butil ay nakakakuha ng isang gatas na kulay, habang nagsisimula ang proseso ng pagtitiwalag ng mga butil ng almirol sa endosperm. Maberde ang shell. Ang nilalaman ng kahalumigmigan ng butil ay nabawasan sa 70-75%, ang libreng kahalumigmigan ay naglalaman ng 3-4 beses na higit pa kaysa sa nakatali, ang tuyong bagay sa pagtatapos ng yugto ay nag-iipon ng mga 10% ng bigat ng isang hinog na butil. Ang tagal ng yugto ay 6-7 araw.
Yugto ng gatas- ang butil ay may pare-pareho ng isang gatas na puting masa, ang shell ay berde. Ang kahalumigmigan ng butil sa pagtatapos ng yugto ay bumaba sa 50%, ang ratio ng libreng tubig sa nakatali na tubig ay humigit-kumulang 1.5:1. Ang dami ng tubig sa 1000 hilaw na butil ay nananatiling humigit-kumulang pare-pareho. Sa yugtong ito, ang tuyong bagay ay masinsinang naipon, ang halaga nito ay halos 50% ng bigat ng isang mature na binhi. Ang tagal ng yugto ay 7-10 araw, minsan 10-15 araw.
Malagkit na yugto ng pagkahinog- ang endosperm ay nakakakuha ng pare-pareho ng isang kuwarta; kapag durog, ang mga hibla ay umaabot. Sa shell, unti-unting nawawala ang chlorophyll (nag-iingat sa uka). Ang kahalumigmigan ng butil ay nabawasan sa 35-42%, ang ratio ng libreng tubig sa nakatali na tubig ay 1:1. Ang nilalaman ng dry matter ay umabot sa 85-90% ng maximum. Ang tagal ng yugto ay 4-5 araw.
Waxy phase- ang endosperm ay nagiging waxy, nababanat. Ang mga shell ay nagiging dilaw. Ang chlorophyll sa uka ay nawawala. Ang dami ng tubig ay nabawasan sa 30%. Ang butil ay umabot sa pinakamataas na dami nito. Sa simula ng yugto, ang isang bahagyang pagtaas sa tuyong bagay sa butil ay nagpapatuloy, at sa pagtatapos ay ganap itong huminto. Ang tagal ng yugto ay 3-6 na araw.
- ang endosperm ay nagiging matigas, pulbos o malasalamin kapag nabasag. Ang shell ay tumatagal din sa isang siksik na parang balat na hitsura. Ang kulay ay tipikal para sa kultura at pagkakaiba-iba na ito. Ang tubig ay nakapaloob, depende sa zone at mga kondisyon, 8-22%, kabilang ang 1-8% sa libreng estado. Ang tagal ng yugto ay 3-5 araw, at pagkatapos ay magsisimula ang isang unti-unting proseso ng pagkawala ng sangkap (pag-expire, atbp.).
Ang tagal ng bawat panahon at yugto ay tinutukoy hindi lamang ng mga katangian ng mga species, kundi pati na rin ng mga kondisyon kung saan nagaganap ang pag-unlad ng binhi. Kapaligiran ay maaaring baguhin hindi lamang ang tagal ng isang panahon o yugto, kundi pati na rin ang kanilang kalikasan (ang mga proseso ng pisyolohikal ay maaaring magpatuloy nang masinsinan, o maaaring higit na mapigilan), na nakakaapekto sa paghahasik at pagbubunga ng mga katangian ng mga buto.
Kung ang panahon ay mainit at tuyo sa panahon ng pagbuo ng mga buto o ang lupa ay hindi sapat na basa, iyon ay, ang butil ay nahuhulog sa ilalim piyus o makunan, pagkatapos ay ang tagal ng panahon ay nabawasan, ang mga buto ay walang oras upang maabot ang kanilang normal na haba at pinaikli (isang napakabihirang pangyayari).
Sa ilang mga kaso, ang proseso ng pagsugpo sa halaman at buto ay maaaring pumunta pa (sa mataas na temperatura at kakulangan ng kahalumigmigan): ang matinding pag-aalis ng tubig ng mga buto ay nangyayari, normal. pisyolohikal na estado mga cell, biochemical na proseso sa pagbabago ng binhi. Ang resulta ay maliliit na buto na may maliit na timbang na 1000 butil, kadalasang may mataas na nilalaman ng nitrogenous compounds.
Ang mahalumigmig na panahon na may kanais-nais na temperatura, ang pagkakaloob ng mga sustansya ay nakakatulong sa pagpapahaba ng panahon ng pagbuo at pagbuo ng mahabang mga buto, na, sa ilalim ng kanais-nais na kasunod na mga kondisyon, ay nagiging malalaking buto.
Ang bigat at sukat ng mga buto ay nakasalalay sa mga kondisyon sa panahon ng pagpuno ng mga buto. Sa ilalim ng normal na kondisyon ng nutrisyon, supply ng tubig at kawalan ng pisikal na pagpapatayo ng mga buto, ang proseso ng pagpuno ay nagpapatuloy sa mas mahabang panahon at maraming organikong bagay. Ang mga buto sa ilalim ng gayong mga kondisyon ay nakakakuha ng isang malaking timbang, sukat, makinis na ibabaw, maliwanag, sariwang kulay, mayroon silang mataas na paghahasik at pagbubunga ng mga katangian.
Sa tag-ulan, ang pag-load ay naantala, ang mga sintetikong proseso ay humina, ang komposisyong kemikal dahil ang ilang mga sangkap ay hindi nagiging mga huling produkto. Ang ganitong mga buto ay nabawasan ang mga katangian ng ani, may mahabang panahon ng pagkahinog pagkatapos ng ani, at hindi maganda ang pag-imbak.
Ang isang mataas na temperatura na may sapat na kumpletong supply ng tubig ay nagpapaikli sa panahon ng pagpuno at nagpapabilis sa bilis ng mga proseso ng biochemical. Ang mga buto ay may mataas na kalidad. Kung ang suplay ng tubig ay hindi sapat, kung gayon, dahil sa pagpapaikli ng panahong ito, ang mga buto ay maaaring maging mahina sa iba't ibang antas. Gayunpaman, ang kahinaan na ito ay nakakaapekto sa kalidad ng mga buto nang hindi gaanong negatibo kaysa sa kahinaan na lumitaw sa panahon ng kanilang pagbuo, kapag hindi kanais-nais na mga kondisyon ay makikita rin sa pagbuo ng embryo.
Ang mga kondisyon na nabubuo sa panahon ng pagkahinog ng binhi ay nakakaapekto sa kanilang kalidad na mas mababa kaysa sa mga kondisyon ng mga nakaraang panahon, ngunit sila ay mahalaga din para sa pagkuha ng mataas na kalidad na mga buto. Sa panahong ito, dapat magkaroon ng pare-pareho, pare-parehong pagpapatuyo ng mga buto, na nag-aambag sa pagbabago ng mga reserbang sustansya sa mga huling anyo. Ang tagtuyot sa yugto ng waxy ripeness, kung nagiging sanhi ito ng mabilis na pagpapatayo ng mga buto, ay humahantong sa isang pagtaas ng nilalaman ng madaling mobile na carbohydrates (asukal, atbp.), na walang oras upang maging almirol. Ang ganitong mga buto ay may mataas na katangian ng paghahasik, lalo na ang mataas na enerhiya ng pagtubo, ngunit nangangailangan ng espesyal na pansin sa panahon ng imbakan. Ang isang tumaas na nilalaman ng mga asukal, kahit na may bahagyang pagtaas sa halumigmig, ay maaaring maging sanhi ng masinsinang paghinga, at kalaunan ay makapinsala sa mga buto.
Ang maulan at malamig na panahon sa panahon ng ripening ay nagpapabagal sa prosesong ito, at ang mga buto ay nakuha na may mahinang mga katangian ng paghahasik at mababang pagtubo. Malamig ngunit tuyong panahon, bagaman nagdudulot ito ng pagpapahaba ng panahon, ngunit ang mga buto ay may kasiya-siyang kalidad.
Ang mga itinuturing na panahon ng pag-unlad ng binhi ay nauugnay sa mga pananim na cereal, ngunit ganap na naaangkop ang mga ito sa iba pang mga pananim, kahit na ang ilang mga yugto ay maaaring magkakaiba.
Pinag-aralan ni V. A. Vishnevsky nang detalyado ang proseso ng pag-unlad ng mga buto ng lupine at itinatag anim na yugto ng pagkahinog: A) ang mga cotyledon ay madilim na berde, ang ugat ng mikrobyo ay berde; b) ang mga cotyledon ay berde, ang simula ng pagpaputi ng ugat ng embryo; V) cotyledon light green, kumpletong pagpaputi ng ugat ng embryo; G) maputi-puti ang mga cotyledon, maagang pagdidilaw ng ugat ng mikrobyo; e) dilaw na cotyledon, dilaw na ugat ng embryo; e) ang mga cotyledon ay dilaw, ang ugat ng embryo ay mapusyaw na dilaw. Ayon sa may-akda, ang panahon ng pagpuno ay nagtatapos sa yugto ng kumpletong pag-yellowing ng ugat ng embryo, kapag ang moisture content ng mga buto ay naging mas mababa sa 50% at ang daloy ng mga plastic substance sa mga buto ay huminto. Ang ganitong paghahati sa mga yugto ng mga panahon ng pagpuno at pagkahinog ay posible para sa iba pang mga munggo, bagaman magkakaroon ng ilang mga pagkakaiba.
Ang proseso ng pagbuo ng mga buto ng mirasol ay makabuluhang naiiba mula sa proseso ng pag-unlad ng mga caryopses. Ayon sa pamamaraan ng V. K. Morozov para sa mirasol itinakda ang mga sumusunod na yugto:
Ang yugto ng pagbuo ng dami ng achene(pericarp) ay nagsisimula nang matagal bago ang pamumulaklak at nagtatapos 6-14 na araw pagkatapos ng pagpapabunga. Ang pericarp ng achene ay lumalaki sa haba para sa mga 6 na araw pagkatapos ng pagpapabunga, at sa lapad at kapal - 8-14 na araw.
Ang yugto ng pagbuo ng dami ng nucleus nagsisimula pagkatapos ng pagpapabunga. Ang kapansin-pansing paglaki sa lahat ng tatlong dimensyon ay magsisimula pagkatapos ng ikaapat na araw at magtatapos sa ika-12-14 na araw.
Yugto ng pagpuno nagsisimula sa dulo ng nauna, at nagtatapos kapag ang supply ng tuyong bagay at ang akumulasyon ng taba sa achene ay tumigil. Ito ay kadalasang nangyayari kapag ang moisture content ng achenes ay bumaba sa 38-40%.
SA ripening phase ang proseso ng pagpapatayo, pag-alis ng kahalumigmigan. Ang mga buto ay napupunta sa isang estado ng post-harvest ripening.
Sa loob ng yugto ng pagkahinog, nakikilala rin ng may-akda antas ng pagkahinog (paghihinog): pag-aani– ang mga buto ay may moisture content na 18–20%, ekonomiya– halumigmig ng achenes 12–14% at overstay– ang moisture content ng mga buto ay mas mababa sa 12%.
Tulad ng nakikita natin, ang dibisyong ito ng proseso ng pag-unlad ng achenes ay batay sa kanilang kahalumigmigan, at sa unang dalawang yugto lamang ay kinuha ang iba pang mga character.
Posibleng ipagpatuloy ang pagsusuri ng mga yugto ng pag-unlad ng iba pang mga kultura, ngunit lahat ng mga ito ay magpapakita lamang ng kanilang pagtitiyak, at ang pangkalahatang pattern ay nananatiling pareho.
Ang isang buto ay ang mikrobyo ng isang bagong organismo, ito ay isang pagbuo sa tulong ng kung saan ang mga halaman ng binhi ay kumakalat at nagpaparami, pati na rin ang karanasan ng mga masamang kondisyon. Ang buto ay bubuo mula sa ovule, kaya marami sa mga bahagi ng ovule, lumalaki at sumasailalim sa iba't ibang pagbabago, ay nagiging mga bahaging bumubuo buto.
Karaniwan, ang isang binhi ay binubuo ng isang binhing mikrobyo, mga sustansya na naiipon sa mga espesyal na trophic tissues (endosperm, perisperm) o sa mismong embryo, at ang seed coat (scleroderma, testa).
karamihan mahalagang bahagi ang binhi ay ang mikrobyo. Ito ay nangyayari alinman bilang isang resulta ng amphimixis, o sa proseso ng apomixis (T. A. Sautkina, V. D. Poliksenova. Plant reproduction, 2001).
Sa pag-unlad nito, ang embryo ay dumaan sa maraming yugto. Ang pag-unlad nito (embryogenesis) ay nagsisimula, tulad ng nabanggit sa itaas, sa mitotic division ng zygote. Sa panahon ng unang dibisyon ng zygote nucleus, ang isang pahalang na septum ay inilalagay sa dulo ng telophase, na nagreresulta sa pagbuo ng dalawang mga selula - basal at apikal (terminal) (Fig. 20d). Kadalasan, bilang isang resulta ng isang serye ng mga mitotic division, isang suspensyon (suspensor) ay nabuo mula sa basal cell, at ang aktwal na embryo ay nabuo mula sa terminal cell. Gayunpaman, hindi lamang ito ang uri ng embryogenesis. Sa ilang mga halaman, ang mga derivatives ng terminal cell ay bahagi ng embryo, habang sa iba, ang mga derivatives ng terminal cell ay kasangkot sa pagbuo ng suspensyon. Ang isang dulo (basal) ng mga palawit ay nakikipag-ugnay sa lugar ng micropyle na may mga tisyu ng nucellus, at sa dulo ng dulo nito ay mayroong isang pre-embryo (madalas na tinatawag na "embryo"), isang spherical formation na binubuo ng meristematic cells (Larawan 19 a, b). Ang palawit ay gumaganap ng isang trophic function, ang mga cell nito ay madalas na bumubuo ng haustoria, tumagos sa tissue ng nucellus at kinuha mula sa mga tisyu nito sustansya, na kinakailangan para sa normal na pag-unlad ng umuusbong na embryo. Bilang karagdagan, unti-unting inililipat ng suspensyon ang pagbuo ng embryo sa gitna ng embryo sac, kung saan matatagpuan ang karamihan sa mga endosperm nuclei (Larawan 19c).
Sa mga unang yugto ng pag-unlad, ang mga embryo ng dicots at halamang monocot magkaroon ng malinaw na pagkakahawig. Nagsisimula silang bumuo bilang mga simetriko na pormasyon, gayunpaman, sa kurso ng pagkita ng kaibhan, ang mga embryo halamang dicot nagiging monosymmetrical formations, at ang mga embryo ng monocots ay nagiging asymmetric. Ito ay dahil sa lokasyon ng kono ng paglago ng stem, na sa mga dicots ay sumasakop sa isang terminal na posisyon, at sa monocots - lateral.
Habang lumalaki ang spherical pre-embryo, nagsisimula ang pagkakaiba-iba nito. Ang una sa spherical body ng pre-embryo ng mga dicotyledonous na halaman, bilang mga lateral formations, ay inilalagay sa anyo ng mga tubercles ng meristematic tissue ng cotyledon. Sa pagitan ng mga ito ay nakahiwalay na kono ng paglago ng tangkay (tugatog ng shoot). Matapos ang pagbuo ng shoot apex, ang mga dahon ng embryonic ay inilalagay bilang mga lateral outgrowth sa growth cone ng stem, at ang embryonic bud ay nabuo. Kaya, ang mga cotyledon (mga dahon ng cotyledonous) at mga dahon ng mikrobyo ay may magkaibang pinagmulan. Sa monocots, isang cotyledon lamang ang nabuo sa panahon ng pagbuo ng embryo.
Kasunod ng pagkita ng kaibahan ng growth cone ng stem sa tapat na dulo ng preembryo, katabi ng suspension, naghihiwalay ang root growth cone. Dahil sa paghahati ng mga selula nito at paglago ng apical, nabuo ang germinal root. Naiiba ang germinal stem sa pagitan ng germinal root at germinal bud. Kaya, ang mga simulain ng mga vegetative organ ng hinaharap na halaman ay inilatag sa buto: ang germinal root, ang germinal stalk at ang germinal bud.
Ang normal na pagkakaiba-iba ng embryo ay nangyayari lamang kung ang endosperm ay nabuo at normal na umuunlad sa mga unang yugto ng pag-unlad ng binhi. Kung ang pag-unlad ng endosperm sa mga unang yugto ay abnormal o ito ay maagang bumagsak, ang nutrisyon ng embryo ay nabalisa, at ang pagkakaiba nito sa mga germinal na organo ay hindi nangyayari. Ang hindi pangkaraniwang bagay na ito ay sinusunod sa mga orchid, sa mga mature na buto kung saan nabuo ang isang hindi nakikilalang masa ng mga cell ng mikrobyo - protokorm. Kung ang endosperm ay bumagsak sa mga huling yugto ng pag-unlad, pagkatapos ng pagkita ng kaibhan ng embryo (pamilya Fabaceae - Legumes), walang mga abnormalidad sa pag-unlad ng buto ang naobserbahan.
Ang embryo ay maaaring magkaroon ng ibang hugis at sumakop sa ibang posisyon sa buto. Ang pinakakaraniwan ay isang direktang embryo, kung saan ang mga cones ng paglago ng stem at ugat ay matatagpuan sa parehong axis (Nicotiana - Tobacco). Ang embryo ay maaaring baluktot (Dianthus - Carnation), spirally twisted (Cuscuta - Dodder), hugis-singsing (Agrostemma - Cockle), hugis ng horseshoe (Capsella - Shepherd's Purse).
Ang embryo sa buto ay maaaring sakupin ang isang sentral na posisyon (Euphorbia - Euphorbia), peripheral (Kukol) at lateral (Cereals).
Ang mga reserbang sustansya ay maaaring maipon hindi lamang sa endosperm. Ayon sa lokalisasyon ng mga reserbang nutrients sa mga mature na buto, nahahati sila sa 4 na grupo:
1. Mga buto na may endosperm.
2. Mga buto na may perisperm.
3. Mga buto na may endosperm at perisperm.
4. Mga buto na walang endosperm.
Ang endosperm ay isang polyploid reserve nutrient tissue na nabuo lamang bilang resulta ng proseso ng fertilization ng polar nuclei o ang central cell ng embryo sac. Sa mga unang yugto ng pag-unlad, ang endosperm ay may mataas na aktibidad ng physiological at biochemical, ngunit ang synthesis ng mga reserbang nutrients sa yugtong ito ay hindi nangyayari dito. Unti-unti, habang naipon ang mga reserbang nutrients, bumababa ang physiological at biochemical na aktibidad ng endosperm. Sa endosperm, ang mga taba, almirol at mga protina ay naipon bilang pangunahing reserbang sustansya, gayunpaman, bilang panuntunan, sa endosperm iba't ibang halaman pinangungunahan ng isang uri ng sustansya. Sa mga cereal, nangingibabaw ang starch, sa sunflower (Helianthus annuus), flax (Linum usitatissimum), at peanut (Arachis hypogea) fats ang nangingibabaw. Bilang karagdagan sa mga pangunahing reserbang nutrients, ang iba pang mga sangkap ay natagpuan sa endosperm ng iba't ibang mga halaman: amino acids (17 amino acids ay ibinibigay para sa mais at trigo), bitamina A, B (1,2,6), C, E, H , PP, at isang bilang ng mga enzyme. Gayunpaman, dahil ang mga mature na buto ay naglalaman ng hindi hihigit sa 14% na tubig, ang endosperm ay physiologically "dead tissue". Ang pagkakapare-pareho ng endosperm ay maaaring mas marami o mas kaunting likido o solid. Sa matigas na endosperm, ang ibabaw ay karaniwang makinis, ngunit sa mga kinatawan ng ilang mga pamilya (Juglandaceae - Walnut, Aristolochiaceae - Guernsey, Arecaceae - Palms) ito ay bumubuo ng mga fold, na pinatataas ang lugar ng pakikipag-ugnay sa embryo. Ang endosperm na ito ay tinatawag na ruminated. Ang isang malakas na nabuong endosperm ay nabuo sa mga buto ng cereal (family Gramineae), lilies (family Liliaceae), nightshade (family Solanaceae), at payong (family Umbellifereae). Ang anyo ng endosperm sa mga buto ng iba't ibang Umbelliferae ay magkakaiba kaya ang karakter na ito ay ginagamit sa taxonomy kapag kinikilala ang mga species bilang isa sa pinakamahalagang diagnostic character.
Ang mga kinatawan ng mga pamilyang Clove (Caryophyllaceae), Chenopodiaceae, at Pepper (Piperaceae) ay walang endosperm sa mature na buto, at ang perisperm ay ang reserbang nutrient tissue. Ito ay nabuo mula sa nucellus at, hindi katulad ng endosperm, ay isang diploid tissue.
Sa mga legume, Asteraceae, kalabasa (Cucurbitaceae) sa mga mature na buto, ang endosperm ay alinman sa ganap na wala o isang maliit na halaga nito ay nananatili sa lugar ng germinal root. Sa mga kinatawan ng mga pamilyang ito, ang mga reserbang sustansya ay idineposito sa mismong embryo, sa mga sobrang hypertrophied na cotyledon nito. Walang endosperm sa mga buto ng orchid (Orchidaceae).
Medyo bihira, dalawang uri ng nutrient tissue ang nabuo sa mga buto - endosperm at perisperm (pamilya Nymphaeaceae - Water lilies).
Ang buto ay natatakpan ng mas marami o hindi gaanong siksik na seed coat. Ang seed coat ay pangunahing bubuo mula sa mga integument ng ovule. Ang isa o parehong mga integument ay maaaring makilahok sa pagbuo ng seed coat, kung minsan ang mga nucellus tissue ay nakikibahagi din sa pagbuo nito.
Pinoprotektahan ng seed coat ang seed embryo mula sa masamang epekto. Sa ilang mga halaman, ito ay natatakpan ng isang cuticle o isang layer ng wax, na ginagawang mas matibay. Ang seed coat ay may isang bilang ng mga tampok na istruktura, ang mga ito ay partikular sa species, mahalaga para sa taxonomy, at nagpapakita ng mga tampok ng pamamahagi ng binhi. Ang mga buto na nabubuo sa mga bukas na prutas ay kadalasang nagkakaroon ng proteksiyon na layer ng mga sclerotesta na selula, ang sclerotesta, sa seed coat. Sa mga buto na ikinakalat ng mga ibon at mammal, ang panlabas na layer ng seed coat ay nagiging makatas at mataba, ito ay tinatawag na sarcotesta. Ang mucilaginous epidermis ng seed coat ay tinatawag na myxotesta. Itinataguyod nito ang pagsipsip ng kahalumigmigan, ikinakabit ang mga buto sa lupa.
Sa ilang mga halaman, ang isang mataba na pormasyon ay bumubuo sa mga buto, bahagyang o ganap na sumasakop sa buto - ang punla, o aryllus. Karaniwan itong maliwanag na kulay, ang mga tisyu nito ay naglalaman ng mga asukal, langis, protina. Ang Arillus ay nagtataguyod ng pagkalat ng mga buto ng mga ibon (Euonimus - Euonymus), mga langgam (Chelidonium - Celandine, Viola - Violet), hangin (Corydalis - Corydalis), tubig (Nymphaeae - Water lily). Sa milkweed (Euphorbia), istoda (Polygala), boxwood (Buxus), isang maliit na paglaki ay nabuo mula sa integument sa lugar ng micropyle - caruncle, na nag-aambag din sa pagkalat ng mga buto.
Bilang karagdagan sa mga partikular na tampok, ang seed coat ng iba't ibang mga halaman ay nailalarawan din ng isang bilang ng karaniwang mga tampok mga gusali. Sa ibabaw ng seed coat, ang isang peklat ng binhi ay malinaw na nakikita - isang bakas na natitira sa lugar ng pagkakabit ng buto sa tangkay ng binhi (funicular). Sa seed coat ay may seminiferous entrance, ang dating micropyle. Sa pamamagitan nito, pumapasok ang tubig sa tumutubo na binhi, at sa pamamagitan nito, kapag tumubo ang binhi, lumalabas ang ugat ng sibol. Karaniwan, ang isang espesyal na pampalapot ay makikita din sa balat ng buto - ang seed suture.
katangian na tampok seed coat ang kulay nito. Ito ay ibinibigay ng pagkakaroon ng iba't ibang mga pigment. Kadalasan, ang mga pigmented na lugar ay kahalili ng mga di-pigmented, na bumubuo ng isang pattern ng katangian (mga buto ng castor). Ang kulay ng seed coat ay nakakatulong din sa dispersal ng binhi.
Ang nabuong mature na binhi ay nailalarawan sa mababang kahalumigmigan at kaunting aktibidad ng mga hormone at enzyme. Sa ganitong estado, ang mga buto ay kayang tiisin ang masamang kondisyon sa kapaligiran at maaaring mapanatili ang kakayahang tumubo nang mahabang panahon. Ang estado na ito ay tinatawag na physiological dormancy ng mga buto.
Ang mga buto ng angiosperms ay magkakaiba sa laki, hugis, masa. Kadalasan ang mga ito ay maliit (hanggang sa 1 cm ang haba), ngunit kung minsan ay umaabot ng ilang sampu-sampung sentimetro ang haba. Sa tropical creeper entada (Entada family Legumes), ang mga buto ay hugis puso at bahagyang mas maliit kaysa sa dessert plate. Ang mga buto ng orkid ay maalikabok na mga 3-5 microns ang lapad. May mga species ng halaman na may bihirang pare-pareho ang hugis at masa. Kaya, ang mga buto ng puno ng carob (Ceratonia siliqua ng pamilyang Cesalpiniaceae) na tumitimbang ng 0.2 g ay kinuha bilang sukatan ng masa sa alahas at tinawag na "carat" (mula sa Arabic karat - butil). Gayunpaman, kadalasan ang hugis at bigat ng mga buto sa parehong halaman ay malaki ang pagkakaiba-iba. Ang phenomenon na ito ay tinatawag na heterospermia (heterospermia) (Calendula officinale - Medicinal marigolds). Ang mga buto ng naturang mga halaman ay naiiba din sa pagtubo, na nagsisiguro sa paglikha ng isang seed bank sa lupa at nag-aambag sa pangmatagalang pangangalaga ng mga species sa isang partikular na lugar.
Kaya, ang pagbuo sa proseso ng ebolusyon ng binhi bilang isang yunit ng pagpaparami at pagpapakalat ng halaman ay lumikha ng isang bilang ng mga pakinabang na nag-ambag sa makabuluhang pagkalat ng mga angiosperms. Kasama sa mga benepisyong ito ang:
Ang angiosperms ay hindi nangangailangan ng isang drop-liquid medium para sa sekswal na proseso at samakatuwid ay mas mahusay na inangkop sa mga kondisyon ng pagkakaroon.
Pinoprotektahan ng seed coat ang embryo.
Ang binhi ay naglalaman ng mga reserbang sustansya na kinakailangan para sa pagbuo ng punla.
Ang mga buto ay kadalasang may mga adaptasyon para sa dispersal.
Ang mga buto ay maaaring manatiling tulog nang mahabang panahon at matiis ang masamang kondisyon.
Ang mga buto ay nabuo sa pamamagitan ng kumbinasyon ng mga asexual at sekswal na proseso, na nagbibigay sa mga species ng mga benepisyo na nauugnay sa genetic variability.
Pagtatapos ng trabaho -
Ang paksang ito ay kabilang sa:
BULAKLAK BILANG ESPESYAL NA REPRODUCTIVE ORGAN
Pag-unlad mas matataas na halaman bryophyte lycopsform horsetail-like fern-like gymnosperms and angiosperms are ... Mga tampok ng pag-unlad ng vegetative sphere, ang pagbuo at istraktura ng vegetative ... Salamat sa vegetative organs na gumaganap ng mga tungkulin ng nutrisyon ng lupa at hangin, transpiration, gas exchange ...
Kung kailangan mo ng karagdagang materyal sa paksang ito, o hindi mo nakita ang iyong hinahanap, inirerekumenda namin ang paggamit ng paghahanap sa aming database ng mga gawa:
Ano ang gagawin natin sa natanggap na materyal:
Kung ang materyal na ito ay naging kapaki-pakinabang para sa iyo, maaari mo itong i-save sa iyong pahina sa mga social network: