Возникновение одноклеточных и многоклеточных водорослей, возникновение фотосинтеза: выход растений на сушу (псилофиты, мхи, папоротники, голосеменные, покрытосеменные).
Развитие растительного мира совершалось в 2 этапа и связано с появлением низших и высших растений. По новой систематике к низшим относят водоросли (а раньше относили бактерии, грибы и лишайники. Теперь они выделены в самостоятельные царства), а к высшим - мхи, папоротникообразные, голосеменные и покрытосеменные.
Возникновение цветка и плода
Первые животные, эксплуатирующие новые наземные среды обитания, созданные наземными растениями, были потомками морских членистоногих, самых разнообразных из всех групп животных. Членистоногие сегодня включают ракообразных; паукообразные; многоножки и многоножки; и насекомых.
То, что сделало членистоногих идеально подходящими для перехода на землю, было их основным планом тела. Их жесткое внешнее покрытие и тонкие, сочлененные придатки оставили несколько мягких частей, подверженных воздействию испарительного эффекта сухого воздуха. Ноги также предоставляли членистоногим средствам передвигаться по суше - перемещаться по препятствиям и прокладывать себе путь, несмотря на большее тяготение силы тяжести, которое приходит с живой водой.
В эволюции низших организмов выделяются 2 периода, существенно различающиеся между собой организацией клетки. В течении 1 периода господствовали организмы, сходные с бактериями и сине-зеленые водорослями. Клетки этих жизненных форм не имели типичных органоидов (митохондирий, хлоропластов, аппарата Гольджи и др.).Ядро клетки не было ограничено ядерной мембраной (это прокариотический тип клеточной организации). 2 период был связан с переходом низших растений (водорослей) к автотрофному типу питания и с образованием клетки со всеми типичными органоидами (это эукариотический тип клеточной организации, который сохранился и на последующих ступенях развития растительного и животного мира). Этот период можно назвать периодом господства зеленых водорослей, одноклеточных, колониальных и многоклеточных. Простейшими из многоклеточных являются нитчатые водоросли (улотрикс), которые не имеют никакого ветвления своего тела. Их тело представляет собой длинную цепочку, состоящую из отдельных клеток. Другие же многоклеточные водоросли расчленены большим количеством выростов, поэтому их тело ветвится (у хары, у фукуса).
Первоначальный переход растений из воды в землю происходил только один раз, и из тех самых ранних новаторских наземных приспособлений происходили все другие линии наземных растений подряд. Одна из причин этого заключалась в том, что растения в их современной форме - как сосудистые, фоосинтезирующие организмы - существовали в океане только в самых примитивных формах водорослей непосредственно перед их перемещением в сушу. Ранние растения развивались на разных этапах адаптации к суше; это привело к увеличению диверсификации с течением времени в четырех основных группах существующих растений.
Многоклеточные водоросли в связи с их автотрофной (фотосинтетичесой) деятельностью развивались в направлении увеличения поверхности тела для лучшего поглощения питательных веществ из водной среды и солнечной энергии. У водорослей появилась более прогрессивная форма размножения - половое размножение, при котором начало новому поколению дает диплоидная (2н) зигота, сочетающая в себе наследственность 2-х родительских форм.
В отличие от растений, членистоногие уже были хорошо установлены в океанах, озерах и ручьях до их миграции на землю. Представители различных групп членистоногих сделали свои. Переходы на землю независимо друг от друга, и каждая группа разработала свои собственные уникальные решения для проживания в сухой среде обитания. Этот переход происходил на протяжении примерно 160 миллионов лет, от позднего ордови-циана до позднего карбонового периода. В то время как первые переходящие членистоногие предшествовали первым переходящим позвоночным на землю, по Каменноугольному периоду, обе группы процветали в обширных тропических болотах и лесах, связанных с месторождениями ископаемого угля.
2 эволюционный этап развития растений необходимо связывать с постепенным переходом их от водного образа жизни к наземному. Первичным наземным организмами оказались псилофиты, которые сохранились в виде ископаемых остатков в силурийских и девонских отложениях. Строение этих растений более сложное по сравнению с водорослями: а) они имели специальные органы прикрепления к субстрату - ризоиды; б) стеблевидные органы с древесиной, окруженной лубом; в) зачатки проводящих тканей; г) эпидермис с устьицами.
Существование многоножек, многоножек, насекомых и пауков наряду с первыми тетраподами обеспечивало устойчивую экосистему, в которой большинство животных были хищниками или мусорщиками. В летописи окаменелостей нет никаких доказательств для еды живых растений либо членистоногими, либо позвоночными до позднекаменноугольной эпохи и пермского периода. До этого времени предполагается, что членистоногие либо были питателями детритов на мертвом органическом веществе, например растительными останками, либо хищниками, которые съели других членистоногих, например насекомых.
Начиная с псилофитов, нужно проследить 2 линии эволюции высших растений, одна из которых представлена мохообразными, а вторая - папоротникообразными, голосеменными и покрытосеменными.
Главное, что характеризует мохообразные, это преобладание в цикле их индивидуального развития гаметофита над спорофитом. Гаметофит - это все зеленое растение, способное к самостоятельному питанию. Спорофит представлен коробочкой (кукушкин лен) и полностью зависит в своем питании от гаметофита. Доминирование у мхов влаголюбивого гаметофита в условиях воздушно-наземного образа жизни оказалось нецелеособразным, поэтому мхи стали особой ветвью эволюции высших растений и пока не дали после себя совершенных групп растений. Этому способствовал и тот факт, что гаметофит по сравнению со спорофитом имел обеденную наследственность (гаплоидный (1н) набор хромосом). Эта линия в эволюции высших растений называется гаметофитной.
Только три из четырех групп членистоногих представлены наземными видами во время палеозоя. Сегодня ракообразные, хотя в основном морские, также имеют наземных членов в виде изоподы и некоторых видов раков-отшельников, но ни одна из них не была среди первой линии наземных животных, покидавших океаны во время палеозоя.
Мириапода: многоножки и многоножки. Многоножки и многоножки являются членами группы членистоногих, известных как Мириапода, имя, которое является греческим для «многих ног». Большинство членистоногих конечностей являются бирамальными или двойными разветвленными; они состоят из крепкого, поддерживающего вес члена, который также имеет ветвящуюся, пернатую, жаберную конечность.
Вторая линия эволюции на пути от псилофитов к покрытосеменным является спорофитной, потому что у папоротникообразных, голосеменных и покрытосеменных в цикле индивидуального развития растений доминирует спорофит. Он представляет собой растение с корнем, стеблем, листьями, органами спороношения (у папоротников) или плодоношения (у покрытосеменных). Клетки спорофита имеют диплоидный набор хромосом, т.к. они развиваются из диплоидной зиготы. Гаметофит сильно редуцирован и приспособлен только для образования мужских и женских половых клеток. У цветковых растений женский гаметофит представлен зародышевым мешком, в котором находится яйцеклетка. Мужской гаметофит образуется при проростании пыльцы. Он состоит из одной вегетативной и одной генеративной клеток. При прорастании пыльцы из генеративной клетки возникает 2 спермия. Эти 2 мужские половые клетки участвуют в двойном оплодотворении у покрытосеменных. Оплодотворенная яйцеклетка дает начало новому поколению растения - спорофиту. Прогресс покрытосеменных обусловлен совершенствованием функции размножения.
Многие виды членистоногих потеряли жаберную конечность, оставив только поддерживающую, однообразную ходячую конечность. Первые свидетельства наземных животных найдены в виде следовых ископаемых с поздней ордовикской эпохи. Это доказательство, состоящее из сохранившихся нор, свидетельствует о том, что к этому времени многоножки эволюционировали и стали важной частью эволюционирующей наземной экосистемы. Первые окончательные окаменелости тела многоножек найдены в позднелурских и раннедевонских отложениях в Шотландии.
Относительная до позднего девона летопись Мириаподы скудная, но сороконожие образцы были найдены в ископаемых несущих породах, известных из фауны Гилбоа в Нью-Йорке и других североамериканских и европейских объектах. Несмотря на небольшой разрыв в летописи окаменелостей, Мириапода продолжал процветать и хорошо диверсифицироваться в каменноугольный период.
|
Выводы:
1. Изучение геологического прошлого Земли, строения и состава ядра и всех оболочек, полеты космических аппаратов на Луну, Венеру, изучение звезд приближает человека к познанию этапов развития нашей планеты и жизни на ней.2. Процесс эволюции носил естественный характер.
3. Растительный мир многообразен, это многообразие есть результат его развития в течение длительного времени. Причина его развития - не божественная сила, - а изменение и усложнение строения растений под влиянием изменяющихся условий среды обитания.
Научные доказательства: клеточное строение растений, начало развития из одной оплодотворенной клетки, необходимость воды для жизненных процессов, нахождение отпечатков различных растений, наличие "живых" ископаемых, вымирание некоторых видов и образование новых.
Они были географически распространены, и их окаменелости были найдены в скалах, которые варьируются в возрасте от позднего силура до раннего пермского периода, в Европе и Северной Америке. В отличие от знакомых многоножек, чьи сегментированные бронежилеты обычно округлые или цилиндрические, артроплеуриды имели широкие плоские спинки, которые спрятали их многочисленные ноги.
В Австралии был обнаружен хотя бы один род. Трек в Нью-Мексико напоминает две параллельные шины. Треки предполагают, что этот экземпляр был длиной не менее восьми футов - настоящая монстра-многоножка своего времени или любого времени. Атмосферный кислород и гигантизм у палеозойских членистоногих.
] [ Украинский язык ] [ Белорусский язык ] [ Русская литература ] [ Белорусская литература ] [ Украинская литература ] [ Основы здоровья ] [ Зарубежная литература ] [ Природоведение ] [ Человек, Общество, Государство ] [ Другие учебники ]
Слизняки или слизистые грибы
Атмосфера Земли претерпела много радикальных сдвигов в своей основной химии в эпоху палеозоя. Наиболее значимо для наземных животных, геохимические исследования осадочных пород и молекул кислорода в воздухе подтверждают заметное увеличение количества атмосферного кислорода в позднем палеозое, за которым следует резкое снижение. Этот импульс кислорода сопровождался столь же резкими сдвигами в количестве углекислого газа в воздухе. По сравнению с современными уровнями кислорода в 21% атмосферы концентрация свободного кислорода в позднем палеозое достигла максимума в карбоне на 35%, а затем упала до 15% к концу перми.
Основные этапы и направления эволюции растительного мира. До конца силурийского периода растения были представлены многоклеточными водорослями, которые либо плавали в воде, либо вели прикрепленный образ жизни. Многоклеточные водоросли явились исходной ветвью для наземных листостебельных растений. В конце силурийского периода палеозойской эры в связи с интенсивными горообразовательными процессами, сокращением площади морей часть водорослей, оказавшись в новых условиях среды (в мелких водоемах и на суше), погибла. Другая часть в результате разнонаправленной изменчивости и адаптации к наземной среде приобрела признаки, способствовавшие выживанию в новых условиях. Такими признаками у первых наземных растений - риниофитов - являются дифференциация тканей на покровные, механические и проводящие и наличие оболочки у спор. Выход растений на сушу был подготовлен деятельностью бактерий и цианобактерий, которые при взаимодействии с минеральными веществами образовали на поверхности суши почвенный субстрат.
Хвощеобразные, или Эквезитофиты
Большая часть этого расширения атмосферного кислорода объясняется ростом растений сосудистых растений, фотосинтез которых вызывает кислород как побочный продукт. Изменения в составе атмосферы повлияли на эволюцию первых животных. Во время перехода от воды к земле первые земные жители столкнулись с сложными климатическими условиями, особенно в отношении способности животных разрабатывать эффективные физиологические средства для извлечения кислорода из воздуха. Членистоногие из воды дышали с помощью жабр.
В девонском периоде на смену риниофитам пришли плауны, хвощи и папоротники, также размножающиеся спорами и предпочитающие влажную среду. Их появление сопровождалось возникновением вегетативных органов, что повышало эффективность функционирования отдельных частей растений и обеспечивало их деятельность как целостной системы.
Членистоногие на суше развивали разнообразные уникальные респираторные стратегии. Насекомые развили особенно эффективный метод дыхания без легких. Они обошли свою систему кровообращения и прямо влили свои ткани кислородом через сеть крошечных ветвящихся трубок, которые бежали от поверхности их жесткой внешней оболочки. респираторная система насекомых эффективна только при небольшом количестве кислорода в воздухе, что является критическим требованием на ранних стадиях эволюции насекомых, когда имелось явно меньшее количество кислорода.
В каменноугольном периоде (карбоне) появляются первые голосеменные, возникшие от древних семенных папоротников. Возникновение семенных растений имело большое значение для дальнейшего развития растительного мира, так как половой процесс стал независимым от наличия капельножидкой среды. Возникшие семенные растения могли обитать в более сухом климате. В пермском периоде климат во многих районах Земли стал более сухим и холодным, древовидные споровые растения, достигшие своего расцвета в карбоне, вымирают. В этот же период начинается расцвет голосеменных, господствовавших в мезозойскую эру. Эволюция высших наземных растений пошла по пути все большей редукции гапло идного поколения (гаметофита) и преобладания диплоидного поколения (спорофита).
К концу девонского периода и к каменноугольному, однако, уровни кислорода на планете быстро росли, поскольку земные планы распространялись повсюду. В летописи окаменелостей показано, что наибольшие насекомые всех времен жили во время позднего палеозоя, особенно в тропических угольных лесах.
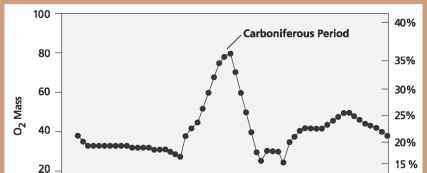
Уровни атмосферного кислорода за последние 550 миллионов лет, что свидетельствует о резком всплеске в период карбона. Тараканы, стрекозы, подмышки и другие насекомые росли до беспрецедентных размеров. Одним из традиционных объяснений эволюции более крупных и крупных насекомых была защита от хищников. Большинство существ того времени были активными хищниками, и одна явная защита от того, чтобы быть съеденным, была для естественного отбора в пользу более крупных членов видов насекомых, которые могли выжить до репродуктивной зрелости.
В меловой период произошел следующий крупный шаг в эволюции ра стений - появились покрытосеменные растения. Первые представители этой группы растений были кустарниками или низкорослыми деревьями с мелкими листьями. Затем довольно быстро покрытосеменные достигли огромного разнообразия форм со значительными размерами и крупными листьями.
Имеет две семядоли
Другим фактором, который способствовал гигантизму у насекомых, была респираторная система насекомых. В то время как насекомые действительно могут выжить при небольшой концентрации кислорода в атмосфере, их система рассеивания кислорода непосредственно в их тканях также допускала впечатляющий рост, когда в позднем палеозое было избыточное количество кислорода. Большие насекомые имели большую площадь поверхности, чтобы поглощать кислород, и эти.
Примеры закрытых тестов
Более крупные насекомые, очевидно, передавали эту черту новым поколениям. В результате, спустя много миллионов лет, был мир, населенный необычно крупными насекомыми, до тех пор, пока не были достигнуты верхние пределы размера этого типа роста. Концентрация кислорода в атмосфере в позднем палеозое также имела последствия для развития полета насекомых. В частности, больше кислорода в воздухе делало воздух более плотным. Это облегчило для крылатых насекомых, чтобы они поднялись, когда они бежали. Меньшие крылья могли бы нести больше веса.
Приобретение различных приспособлений для опыления цветков и рас пространения плодов и семян позволило покрытосеменным занять в кай нозое господствующее положение в растительном мире.
Таким образом, основными особенностями эволюции растительного мира были:
- постепенный переход к доминирующему положению спорофита над
гаметофитом в цикле развития; - выход на сушу, дифференциация тела на органы (корень, стебель, лист)
и дифференциация тканей (проводящие, механические, покровные); - переход от наружного оплодотворения к внутреннему; возникнове
ние цветка и двойного оплодотворения; - возникновение семян, содержащих запах питательных веществ и за
щищенных от воздействия неблагоприятных условий среды семенными
покровами (и стенками околоплодника у покрытосеменных);
- совершенствование органов размножения и перекрестного оплодо
творения у покрытосеменных параллельно с эволюцией насекомых; - возникновение разнообразных способов распространения плодов и
семян.
Основные этапы и направления эволюции животного мира. История эволюции животных изучена наиболее полно в связи с тем, что многие из них имеют скелет и поэтому лучше сохраняются в окаменелых остатках.
Успех первых летчиков привел к последовательным поколениям специализации в дизайне крыла и эффективности. В значительной степени эта эволюционная веха стала возможной благодаря плотной атмосфере, которая способствовала развитию раннего полета. Если насекомые и другие членистоногие, такие как скорпионы и пауки, были настолько успешны в использовании богатой кислородом окружающей среды, почему тогда не развивались стрекозы размером с самолеты и скорпионы размером с танки? Ответ заключается в том, что существуют пороги, налагаемые законами физики, которые ограничивают размер тела организмов.
Многоклеточные животные происходят от одноклеточных организмов через колониальные формы. Первыми животными были, вероятно, кишеч нополостные. Древние кишечнополостные дали начало плоским червям, которые являются трехслойными животными с двусторонней симметрией.
От древних ресничных червей произошли первые вторичнополостные животные - кольчатые черви. Древние морские многощетинковые, веро ятно, послужили основой для возникновения типов членистоногих, моллюсков и хордовых.
В случае членистоногих размер тела ограничивается самой дыхательной системой, которая позволяет организму расти настолько велика, в первую очередь. Диффузионный тип дыхания, в котором кислород подается в ткани поверхностными трубками, чрезвычайно эффективен у мелких животных. Однако, если виды насекомых растут, эта форма газообмена работает против дальнейшего роста. Палеонтолог Ричард Фортей указывает, что по мере увеличения площади поверхности - внешней оболочки экзоскелета - членистоногих, внутренний объем животного также увеличивается, но в восемь раз.
Самые древние следы животных относятся к докембрию (около 700 млн лет назад). В кембрийском и ордовикском периодах преобладают губки, кишечнополостные, черви, иглокожие, трилобиты, появляются моллюски.
В позднем кембрии возникают бесчелюстные панцирные рыбы, а в девоне - челюстные рыбы. Для большинства из этих животных характерны наличие двусторонней симметрии, третьего зародышевого листка, полости тела, наружного (членистоногие) или внутреннего (ходовые) твердого скелета, прогрессирующая способность к активному передвижению, обособление переднего конца тела с ротовым отверстием и органами чувств, постепенное совершенствование центральной нервной системы.
Существует точка, в которой более крупная версия того же животного просто не сможет поглощать достаточное количество кислорода для кормления его тканей и органов. Второе физическое ограничение также связано с крошечной трахеей или трубками для диффузии воздуха. По мере роста конечностей членистоногих внутренние поверхности воздушных трубок становятся сильнее и жестче. Существует точка, в которой трубам трудно сгибаться, и любые такие воздушные проходы к тканям членистоногих ног заставляют ноги слишком жесткими, чтобы двигаться.
Любая перспектива - невозможность дышать или невозможность ходить - обеспечивает биологические ограничения для эволюции членистоногих неограниченного размера. Процветал в ответ на обилие кислорода в каменноугольной атмосфере. Когда атмосфера Земли изменилась в пермский период, а уровень кислорода снизился до уровней, которые более похожи на сегодняшние, Артроплура была среди многих крупных видов членистоногих, которые вымерли. Независимо от того, был ли низкий уровень кислорода ответственным за вымирание, до конца не понят, хотя это, возможно, и послужило поводом для кончины такого крупного членистоногих.
От первых челюстноротых возникли лучеперые и кистеперые рыбы. Кистеперые имели в плавниках опорные элементы, из которых позже развились конечности наземных позвоночных. Наиболее важные ароморфозы в этой линии эволюции - развитие из жаберных дуг подвижных челюстей (обеспечивали активный захват добычи), развитие из кожных складок плавников, а затем формирование поясов парных грудных и брюшных конечностей (увеличивали маневренность движений в воде). Двоякодышащие и кистеперые рыбы посредством плавательных пузырей, имеющих связь с пищеводом и снабженных системой кровеносных сосудов, могли дышать атмосферным кислородом.
От кистеперых рыб берут начало первые наземные животные - стегоцефалы. Стегоцефалы разделились на несколько групп амфибий, которые достигли расцвета в карбоне. Выход на сушу первых позвоночных животных был обеспечен преобразованием плавников в конечности наземного типа, воздушгных пузырей - в легкие.
От амфибий ведут свое начало истинно наземные животные - рептилии, завоевавшие сушу к концу пермского периода. Освоение суши пресмыкающимися обеспечило наличие сухих ороговевших покровов, внутреннего оплодотворения, большого количества желтка в яйцеклетке, защитных оболочек яиц, предохраняющих эмбрионы от высыхания и других воздействий среды. Среди рептилий выделилась группа динозавров, давшая начало млекопитающим. Первые млекопитающие появились в триасовом периоде мезозойской эры. Позднее, также от одной из ветвей пресмыкающихся, произошли зубатые птицы (археоптерикс), а затем - современные птицы. Для птиц и млекопитающих характерны такие черты, как теплокровность, четырехкамерное сердце, одна дуга аорты (создает полное разделение большого и малого кругов кровообращения), интенсивный обмен веществ - черты, обеспечившие расцвет этих групп организмов.
В конце мезозоя появляются плацентарные млекопитающие, для которых основными прогрессивными особенностями стали появление плаценты и внутриутробного развития плода, вскармливание детенышей молоком, развитая кора головного мозга. В начале кайнозойской эры от насекомоядных обособился отряд приматов, эволюция одной из ветвей которого привела к возникновению человека.
Параллельно эволюции позвоночных шло развитие беспозвоночных животных. Переход от водной к наземной среде обитания осуществился у паукообразных и насекомых с развитием совершенного твердого наружного скелета, членистых конечностей, органов выделения, нервной системы, органов чувств и поведенческих реакций, появлением трахейного и легочного дыхания. Среди моллюсков выход на сушу наблюдался значительно реже и не приводил к такому разнообразию видов, какое наблюдается у насекомых.
Основные особенности эволюции животного мира:
- прогрессивное развитие многоклеточности и, как следствие, специа лизация тканей и всех систем органов;
- свободный образ жизни, который определил выработку различных механизмов поведения, а также относительную независимость онтогенеза от колебаний факторов окружающей среды;
- возникновение твердого скелета: наружного у некоторых беспозво ночных (членистоногие) и внутреннего у хордовых;
- прогрессивное развитие нервной системы, что явилось основой для возникновения условно-рефлекторной деятельности.
Среди основных этапов эволюции растительного мира можно выделить выход на сушу, переход от наружного оплодотворения к внутреннему, возникновение семян и совершенствование способов их распространения; в эволюции животного мира - специализацию тканей и систем органов, возникновение твердого скелета, прогрессивное развитие нервной системы и возможность вести свободный образ жизни.
1. Когда на суше появились растения? Как вы думаете, что должно было предшествовать выходу растений на сушу? 2. Какие ароморфозы дали растениям возможность закрепиться на суше? 3. Какие животные были первыми обитателями суши? Когда они появились? Какие ароморфозы позволили животным освоить сушу? 4. Какая существует связь между завоеванием позвоночными суши и развитием нервной системы? 5. От кого произошли млекопитающие? Какие ароморфозы определили формирование млекопитающих как класса?
Общая биология: Учебное пособие для 11-го класса 11-летней общеобразовательной школы, для базового и повышенного уровней. Н.Д. Лисов, Л.В. Камлюк, Н.А. Лемеза и др. Под ред. Н.Д. Лисова.- Мн.: Беларусь, 2002.- 279 с
Содержание учебника Общая биология: Учебное пособие для 11-го класса:
- § 2. Популяция - структурная единица вида. Характеристика популяции
- § 6. Экосистема. Связи организмов в экосистеме. Биогеоценоз, структура биогеоценоза
- § 7. Движение вещества и энергии в экосистеме. Цепи и сети питания
- § 9. Круговорот веществ и поток энергии в экосистемах. Продуктивность биоценозов
- § 13. Предпосылки возникновения эволюционной теории Ч. Дарвина
- § 14. Общая характеристика эволюционной теории Ч. Дарвина
- § 18. Развитие эволюционной теории в последарвиновский период. Синтетическая теория эволюции
- § 19. Популяция - элементарная единица эволюции. Предпосылки эволюции
- § 27. Развитие представлений о возникновении жизни. Гипотезы происхождения жизни на Земле
- § 32. Основные этапы эволюции растительного и животного мира
- § 33. Многообразие современного органического мира. Принципы систематики
- § 35. Формирование представлений о происхождении человека. Место человека в зоологической системе
- § 36. Этапы и направления эволюции человека. Предшественники человека. Древнейшие люди
Глава 1. Вид - единица существования живых организмов
Глава 2. Взаимоотношения видов, популяций с окружающей средой. Экосистемы
Глава 3. Формирование эволюционных взглядов
Глава 4. Современные представления об эволюции
Глава 5. Происхождение и развитие жизни на Земле
Глава 6. Происхождение и эволюция человека