Arabic Bulgarian Chinese Croatian Czech Danish Dutch English Estonian Finnish French German Greek Hebrew Hindi Hungarian Icelandic Indonesian Italian Japanese Korean Latvian Lithuanian Malagasy Norwegian Persian Polish Portuguese Romanian Russian Serbian Slovak Slovenian Spanish Swedish Thai Turkish Vietnamese
definition - Доминантность ДоминантностьМатериал из Википедии - свободной энциклопедииДоминантность (доминирование) - форма взаимоотношений между аллелями одного гена, при которой один из них (доминантный) подавляет (маскирует) проявление другого (рецессивного) и таким образом определяет проявление признака как у доминантных гомозигот , так и у гетерозигот .
При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А (сцепленный с Х-хромосомой рецессивный ген), имеют половинное количество нормального фактора свертывания по сранению с гомозиготными по нормальному аллелю людьми, и активность фактора свертывания VIII у них в среднем вдвое ниже, чем у здоровых людей. В то же время у здоровых людей активность этого фактора варьирует от 40 до 300 % по сравнению со средней для популяции. Поэтому наблюдается значительное перекрывание признаков у здоровых и носителей-гетерозигот. При фенилкетонурии (аутосомно-рецессивный признак) гетерозиготы обычно считаются здоровыми, однако активность печёночного фермента фенилаланин-4-гидроксилазы у них вдвое ниже нормы, а содержание фенилаланина в клетках повышено, что, по некоторым данным, приводит к снижению IQ и повышенному риску развития некоторых психотических расстройств.
Неполное доминированиеПри неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если домининатный аллель дает функциональный белок, а рецессивный - дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную - нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
КодоминированиеПри кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно (смешанно). Типичный пример кодоминирования - наследование групп крови системы АВО у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой - рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип.На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов - это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Локальное (внутримолекулярное) доминирование Относительный характер доминированияКак уже отмечалось выше, характер доминирования зависит от уровня анализа признака. Рассмотрим это на примере серповидно-клеточной анемии . Гетерозиготные носители гена гемоглобина S (AS) на уровне моря имеют нормальную форму эритроцитов и нормальную концентрацию гемоглобина в крови (полное доминирование А над S). На больших высотах (более 2,5-3 тыс. м) у гетерозигот концентрация гемоглобина понижена (хотя и намного выше, чем у больных), появляются эритроциты серповидной формы (неполное доминирование А над S). Этот пример показывает, что доминантность может зависеть от условий. Гетерозиготы AS и гомозиготы SS обладают примерно одинаковой устойчивостью к малярии, гомозиготы АА подвержены малярии в большей степени. По данному проявлению ген S доминирует над А. Наконец, в эритроцитах носителей АS в раных количествах присутствуют оба варианта бета-глобиновых цепей - нормальный А и мутантный S (то есть наблюдается кодоминирование).
Эволюция доминантностиНовые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вариант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путем отбора малых мутаций как структурных генов, так и генов-модификаторов.
Так или иначе, степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего темную окраску бабочек березовой пяденицы (Biston betu-laria), которое, по некоторым данным, произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название "индустриальный меланизм) (Н.Н.Иорданский "Эволюция жизни")
Которая может подавляться воздействием доминантного гена и не проявляется в фенотипе . Рецессивный ген способен обеспечить проявление определяемого им признака только в том случае, если находится в паре с соответственным рецессивным геном. Если же он находится в паре с доминантным геном, то он не проявляется, так как доминантный ген подавляет его. Свойства, представленные рецессивными генами, проявляются в фенотипе у потомка лишь в том случае, если у обоих родителей присутствует рецессивный ген.
При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А (сцепленный с Х-хромосомой рецессивный ген), имеют половинное количество нормального фактора свертывания по сравнению с гомозиготными по нормальному аллелю людьми, и активность фактора свертывания VIII у них в среднем вдвое ниже, чем у здоровых людей. В то же время у здоровых людей активность этого фактора варьирует от 40 до 300 % по сравнению со средней для популяции. Поэтому наблюдается значительное перекрывание признаков у здоровых и носителей-гетерозигот. При фенилкетонурии (аутосомно-рецессивный признак) гетерозиготы обычно считаются здоровыми, однако активность печёночного фермента фенилаланин-4-гидроксилазы у них вдвое ниже нормы, а содержание фенилаланина в клетках повышено, что, по некоторым данным, приводит к снижению IQ и повышенному риску развития некоторых психотических расстройств.
Неполное доминированиеПри неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный - дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную - нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
КодоминированиеПри кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно (смешанно). Типичный пример кодоминирования - наследование групп крови системы АВ0 у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой - рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип. На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов - это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Относительный характер доминированияКак уже отмечалось выше, характер доминирования зависит от уровня анализа признака. Рассмотрим это на примере серповидно-клеточной анемии . Гетерозиготные носители гена гемоглобина S (AS) на уровне моря имеют нормальную форму эритроцитов и нормальную концентрацию гемоглобина в крови (полное доминирование А над S). На больших высотах (более 2,5-3 тыс. м) у гетерозигот концентрация гемоглобина понижена (хотя и намного выше, чем у больных), появляются эритроциты серповидной формы (неполное доминирование А над S). Этот пример показывает, что доминантность может зависеть от условий. Гетерозиготы AS и гомозиготы SS обладают примерно одинаковой устойчивостью к малярии, гомозиготы АА подвержены малярии в большей степени. По данному проявлению ген S доминирует над А. Наконец, в эритроцитах носителей АS в равных количествах присутствуют оба варианта бета-глобиновых цепей - нормальный А и мутантный S (то есть наблюдается кодоминирование).
Эволюция доминантностиНовые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вариант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путем отбора малых мутаций как структурных генов, так и генов- модификаторов.
Так или иначе, степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего темную окраску бабочек березовой пяденицы (Biston betu-laria), которое, по некоторым данным, произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название «индустриальный меланизм ») (Н. Н. Иорданский «Эволюция жизни»)
См. также Ссылки- «OMIM Entry - *612349 - PHENYLALANINE HYDROXYLASE; PAH» . OMIM.org.
- Cat Coat Color . Vgl.ucdavis.edu. doi :10.1111/j.1365-2052.2009 . Архивировано
- Carr, Steven M Extensions to Mendelian Analysis . Mun.ca . Memorial University of Newfoundland. Архивировано из первоисточника 19 мая 2012. Проверено 2 ноября 2011.
Wikimedia Foundation . 2010 .
Синонимы : Смотреть что такое "Доминантность" в других словарях:Участие только одного аллеля в определении признака у гетерозиготной особи. Явление Д. открыто ещё в первых классич. опытах Г. Менделя. Доминантные аллели обозначают прописными буквами А, В и т. д. Когда нет доминирования в строгом смысле этого… … Биологический энциклопедический словарь
- (от лат. dominari господствовать) сложившаяся в процессе биотоценогенеза способность некоторых видов занимать в биоценозе главенствующее положеаие и оказывать преобладающее влияние на образование в нем биоценоти ческой среды. Доминантность вида… … Экологический словарь
Современная энциклопедия
- (доминирование) форма взаимоотношений парных (аллельных) генов, при которой один из них доминантный оказывает более сильное влияние на соответствующий признак особи, чем другой рецессивный. Явление доминантности открыто Г. Менделем … Большой Энциклопедический словарь
Сущ., кол во синонимов: 3 властность (13) доминирование (8) понты (9) … Словарь синонимов
доминантность - Властность, склонность и способность занимать господствующее положение. Тематики сексология … Справочник технического переводчика
Доминантность - (доминирование), форма взаимоотношений парных (аллельных) генов, при которой один из них доминантный оказывает более сильное влияние на соответствующий признак особи, чем другой рецессивный. Пример доминантности у человека кареглазость. Сравни… … Иллюстрированный энциклопедический словарь
Неполное доминирование. Всеобщность закона доминирования вскоре после его переоткрытия на основании целого ряда фактов была подвергнута сомнению. Оказалось, что для большого числа признаков у растений и животных характерно промежуточное наследование, или неполное доминирование, в первом поколении. Такое наследование отмечал сам Мендель по некоторым признакам у гороха.
При неполном доминировании гибрид F 1 (Аа) не воспроизводит полностью ни одного из родительских признаков, выражение признака оказывается промежуточным, с большим или меньшим уклонением к доминантному или рецессивному состоянию, но все особи этого поколения одинаковы по фенотипу.
Примером неполного доминирования может быть промежуточная розовая окраска ягоды у гибридов земляники (Fragaria vesca), полученных от скрещивания форм с красной и белой ягодами (рис. 1).

Рисунок 1. Наследование окраски ягоды при неполном доминировании у земляники: АА - красная; аа - белая; Ла - розовая окраска.
При полном доминировании расщепление в F 1 по фенотипу (3:1) не совпадает с расщеплением по генотипу (1:2: 1), так как гетерозигота Аа внешне неотличима от гомозиготы АА. При неполном же доминировании во втором поколении имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота АА отличается от гетерозиготы Аа. Так, в разбираемом примере с земляникой расщепление в F 1 по окраске ягоды 1 красная (АА) :2 розовых (Аа) : 1 белая (аа). В этом случае лишь условно можно назвать красную окраску ягоды доминантной, а белую - рецессивной.
Неполное доминирование проявляется и при скрещивании растений ночной красавицы, имеющей пурпурные цветки (АА), с растением, имеющим белые цветки (аа). Все растения первого поколения будут иметь промежуточную розовую окраску (рис. 2). Это не противоречит правилу единообразия гибридов первого поколения Г. Менделя: ведь действительно в первом поколении все цветки розовые. При скрещивании двух особей ночной красавицы из первого поколения во втором поколении происходит расщепление, но не в соотношении 3:1, а в соотношении 1:2:1, т.е. один цветок белый (аа), два розовых (Аа) и один пурпурный (АА).
Неполное доминирование оказалось широко распространенным явлением, оно наблюдается в наследовании окраски цветка у львиного зева, окраски оперения у кур, шерсти у норок и лошадей, а также многих других морфологических и физиологических признаков у растений, животных и человека.
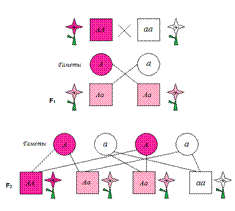
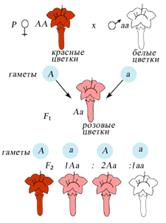
Рисунок 2. Явление неполного доминирования при формировании окраски венчиков цветков ночной красавицы
Сущность доминирования. Доминирование является свойством гена обусловливать развитие признака в гетерозиготном состоянии. Биохимические исследования действия генов, например, у львиного зева (Antirrinum maijus), у которого красная окраска цветка доминирует над светло-красной, также демонстрирует природу доминирования. Биохимический анализ показал, что красный пигмент цианидин отличается от светло-красного пеларгонидина лишним гидрокислом ОН в третьем положении (рис. 3). Каждый из этих двух пигментов синтезируется растением под действием определенного фермента: один из них присоединяет ОН в 4-е положение, благодаря чему вырабатывается пеларгонидин, а второй фермент превращает его в цианидин, присоединяя ОН в 3-е положение, т. е. у гетерозиготного растения в конечном итоге синтезируется цианидин, благодаря чему красная окраска доминирует над светло-красной. Очевидно, можно предполагать, что во всех случаях, когда имеет место подобный биохимический механизм образования пигментов, красная окраска будет доминировать над светло-красной. Действительно, у всех растений, где этот механизм найден, наблюдается и подобный же характер доминирования (стрепто-карпус, тюльпан и др.). Биохимический анализ генных продуктов позволяет предвидеть характер доминирования.
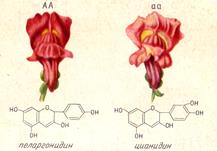 Рисунок 3. Окраска цветка у львиного зева: АА - красная;
Рисунок 3. Окраска цветка у львиного зева: АА - красная;
аа - светло-красная и химические формулы пигментов
В случаях неполного доминирования доминантный ген не до конца подавляет рецессивный ген из аллельной пары. При этом будут возникать промежуточные признаки, и признак у гомозиготных особей будет не таким, как у гетерозиготных. Как выяснилось, это не означает, что рецессивная аллель полностью подавлена и абсолютно не функционирует. Например, при промежуточном наследовании функционируют обе аллели гена. Даже в случае полного доминирования можно найти примеры, свидетельствующие о том, что и рецессивная аллель вырабатывает определенный продукт. Так, у кукурузы (Zea mays) найдены два типа эфиров, которые гидролизуются ферментами, называемыми эстеразами. Наличие этих ферментов определяет пара аллелей. У гетерозиготных растений гидролизуется только один из эфиров под действием фермента, определяемого доминантной аллелью. Биохимический анализ показал, что у гибрида вырабатываются оба фермента, но только доминантная аллель вырабатывает активный фермент, а рецессивная - неактивный. Рецессивная аллель функционирует наравне с доминантной. Возможно, что в результате взаимодействия этих двух ферментов функционирование одного из них, продуцируемого рецессивной аллелью, подавляется. Доминирование проявляется во взаимодействии тех продуктов действия генов - признаков, которые определяются доминантной и рецессивной аллелями.
Управление доминированием. Механизм явления доминирования обусловливает возможность изменения доминирования. Оно изменяется под влиянием внешних условий, в разной степени благоприятствующих или препятствующих развитию одного из пары признаков. И действительно, например, у пшеницы (Triticum) в обычных условиях доминирует нормальный колос, а при коротком световом дне - ветвистый. Но изменение характера доминирования в индивидуальном развитии гибрида не приводит к изменению его генотипа и, следовательно, не изменяет расщепление в его потомстве. Поэтому расщепление в F 2 по форме колоса одинаковое, независимо от того, какой признак доминировал у гибрида F 1 (рис. 4).
Рисунок 4. Доминирование типа колоса у пшеницы в зависимости от длины светового дня: А - нормальный колос; а - ветвистый.
И. В. Мичурин на основании многолетних исследований показал возможность управления доминированием у некоторых гибридов. Для того чтобы получить гибриды с нужными свойствами, он не только тщательно подбирал формы для скрещивания, но и создавал условия, благоприятствующие развитию гибрида в определенном, желательном направлении. Для управления доминированием он разработал метод ментора (воспитателя), заключающийся в прививке гибрида на одну из скрещиваемых форм. Растение, на которое производится прививка, называется подвоем, прививаемую часть растения называют привоем. При срастании тканей растений разных сортов или даже видов и родов под влиянием подвоя в привое могут меняться некоторые физиологические и морфологические особенности. Например, при опылении цветков красноплодной владимирской вишни (Prunus cerasus) пыльцой белоплодной черешни Винклера (P. avium) Мичурин получил гибрид с бледно-розовыми плодами. Черенок гибридного растения, привитый в крону вишни, стал давать темно-розовые плоды. Следовательно, под влиянием прививки изменился характер доминирования. В дальнейшем путем вегетативного размножения этого гибрида был получен сорт Краса Севера. В селекции плодовых, которые размножаются вегетативно и потому при размножении не утрачивают приобретенных в онтогенезе свойств, метод ментора имеет большое значение.
К сожалению, эти опыты И. В. Мичурина не сопровождались цитофизиологическим и биохимическим анализом, и до сих пор не выяснена биохимическая сущность взаимоотношения компонентов прививки.
Изучение наследования различных признаков на разных объектах показало общность основных закономерностей наследования не только для растений, но и для животных – позвоночных и беспозвоночных, а также для человека и микроорганизмов.
Однако, как и всякий закон живой природы, закон расщепления проявляется лишь при определенных условиях. Если эти условия не соблюдаются, то характер расщепления может измениться.
Известны случаи, когда два или более аллелей не проявляют в полной мере доминантность или рецессивность, так что в гетерозиготном состоянии ни один из аллелей не доминирует над другим. Это явление неполового доминирования, или кодоминантность, представляет собой исключение из описанного Менделем правила наследования при моногибридных скрещиваниях. К счастью, Мендель выбрал для своих экспериментов признаки, которым не свойственно неполное доминирование; в противном случае оно могло бы сильно осложнить его первые исследования. Неполное доминирование наблюдается как у растений, так и у животных. В большинстве случаев гетерозиготы обладают фенотипом, промежуточным между фенотипами доминантной и рецессивной гомозигот. Примером служат андалузские куры, полученные в результате скрещивания чистопородных черных и «обрызганных белых» (splashed white) кур. Черное оперение обусловлено наличием аллеля, определяющего синтез черного пигмента меланина. У «обрызганных» кур этот аллель отсутствует. У гетерозигот меланин развивается не в полной мере, создавая лишь голубоватый отлив на оперении.
Поскольку общепринятых символов для обозначения аллелей с неполным доминированием не существует, нам необходимо ввести для генотипов такие символы, чтобы сделать понятными приведенные ниже схемы получения андалузских кур.
Возможны, например, такие обозначения: черные - В, «обрызганные» - b, W, BW или BBW. Результаты скрещивания между гомозиготными черными и «обрызганными» курами представлены в табл. 2. При скрещивании между собой особей F1 отношение фенотипов в F2 отличается от менделевского отношения 3: 1, типичного для моногибридного скрещивания. В этом случае получается отношение 1:2:1, где у половины особей F2 будет такой же генотип, как у F1 (табл. 3). Отношение 1: 2: 1 характерно для результатов скрещиваний при неполном доминировании.
Таблица 2. Скрещивание кур андалузской породы: гибриды F1.
Конец работы -
Эта тема принадлежит разделу:
Хромосомная теория наследственностиВо всех случаях анализ результатов показал что отношение доминантных признаков к рецессивным в поколении F составляло примерно... Приведенный выше пример типичен для всех экспериментов Менделя в которых... На основании этих и аналогичных результатов Мендель сделал следующие выводы...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Возвратное, или анализирующее, скрещивание
Организм из поколения F1, полученного от скрещивания между гомозиготной доминантной и гомозиготной рецессивной особями, гетерозиготен по своему генотипу, но обладает доминантным фенотипом. Для того
Дигибридное скрещивание и закон независимого распределения
Установив возможность предсказывать результаты скрещиваний по одной паре альтернативных признаков, Мендель перешел к изучению наследования двух пар таких признаков. Скрещивания между особями, разли
Краткое изложение сути гипотез Менделя
· Каждый признак данного организма контролируется парой аллелей.
· Если организм содержит два различных аллеля для данного признака, то один из них (доминантный) может проявляться, полност
Хромосомная теория наследственности
К концу XIX в. в результате повышения оптических качеств микроскопов и совершенствования цитологических методов возможно стало наблюдать поведение хромосом в гаметах и зиготах. Еще в 1875 г. Гертви
Сцепление
Все ситуации и примеры, обсуждавшиеся до сих пор, относились к наследованию генов, находящихся в разных хромосомах. Как выяснили цитологи, у человека все соматические клетки содержат по 46 хромосом
Группы сцепления и хромосомы
Генетические исследования, проводившиеся в начале нашего века, в основном были направлены на выяснение роли генов в передаче признаков. Работы Моргана с плодовой мушкой Drosophila melanogaster пока
Гигантские хромосомы и гены
В 1913 г. Стертевант начал свою работу по картированию положения генов в хромосомах дрозофилы, во это было за 21 год до того, как появилась возможность связать различимые в хромосомах структуры с г
Определение пола
Рисунок 1. Хромосомные наборы самца и самки D. melanogaster. Они состоят из четырех пар хромосом (пара I - пол
Летальные гены
Известны случаи, когда один ген может оказывать влияние на несколько признаков, в том числе на жизнеспособность. У человека и других млекопитающих определенный рецессивный ген вызывает образование
Эпистаз
Ген называют эпистатическим (от греч. еpi - над), если его присутствие подавляет эффект какого-либо гена, находящегося в другом локусе. Эпистатические гены иногда называют ингибирующими генами, а т
Изменчивость
Изменчивостью называют всю совокупность различий по тому или иному признаку между организмами, принадлежащими к одной и той же природной популяции или виду. Поразительное морфологическое разнообраз
Дискретная изменчивость
Некоторые признаки в популяции представлены ограниченным числом вариантов. В этих случаях различия между особями четко выражены, а промежуточные формы отсутствуют; к таким признакам относятся, напр
Непрерывная изменчивость
По многим признакам в популяции наблюдается полный ряд переходов от одной крайности к другой без всяких разрывов. Наиболее яркими примерзлая служат такие признаки, как масса (вес), линейные размеры
Влияние среды
Главный фактор, детерминирующий любой фенотипический признак, - это генотип. Генотип организма определяется в момент оплодотворения, но степень последующей экспрессии этого генетического потенциала
Необходимо ясно представлять себе, что взаимодействие между дискретной и непрерывной изменчивостью и средой делает возможным существование двух организмов с идентичным фенотипом. Механизм репликаци
Мутации
Мутацией называют изменение количества или структуры ДНК данного организма. Мутация приводит к изменению генотипа, которое может быть унаследовано клетками, происходящими от мутант- ной клетки в ре
Генные мутации
Внезапные спонтанные изменения фенотипа, которые нельзя связать с обычными генетическими явлениями или микроскопическими данными о наличии хромосомных аберраций, можно объяснить только изменениями
Значение мутаций
Хромосомные и генные мутации оказывают разнообразные воздействия на организм. Во многих случаях эти мутации летальны, так как нарушают развитие; у человека, например, около 20% беременностей заканч
Наследственная изменчивость
Комбинативная изменчивость. Наследственную, или геноти-пическую, изменчивость подразделяют на комбинативную и мутационную.
Комбинативной называют изменчивость, в основе которой лежит об
Дигибридное скрещивание
Сущность дигибридного скрещивания. Организмы различаются по многим генам и, как следствие, по многим признакам. Чтобы одновременно проанализировать наследование нескольких признаков, необходимо изу
Методы генетики
Основным является гибридологический метод - система скрещиваний, позволяющая проследить закономерности наследования признаков в ряду поколений. Впервые разработан и использован Г.
Генетическая символика
Предложена Г. Менделем, используется для записи результатов скрещиваний: Р - родители; F - потомство, число внизу или сразу после буквы указывает на порядковый ном
Закон единообразия гибридов первого поколения, или первый закон Менделя
Успеху работы Менделя способствовал удачный выбор объекта для проведения скрещиваний - различные сорта гороха. Особенности гороха: 1) относительно просто выращивается и имеет короткий период развит
Закон расщепления, или второй закон Менделя
Г. Мендель дал возможность самоопылиться гибридам первого поколения. У полученных таким образом гибридов второго поколения проявился не только доминантный, но и рецессивный признак. Результаты опыт
Закон чистоты гамет
С 1854 года в течение восьми лет Мендель проводил опыты по скрещиванию растений гороха. Им было выявлено, что в результате скрещивания различных сортов гороха друг с другом гибриды первого поколени
Цитологические основы первого и второго законов Менделя
Во времена Менделя строение и развитие половых клеток не было изучено, поэтому его гипотеза чистоты гамет является примером гениального предвидения, которое позже нашло научное подтверждение.
Закон независимого комбинирования (наследования) признаков, или третий закон Менделя
Организмы отличаются друг от друга по многим признакам. Поэтому, установив закономерности наследования одной пары признаков, Г. Мендель перешел к изучению наследования двух (и более) пар альтернати
Цитологические основы третьего закона Менделя
Пусть А - ген, обусловливающий развитие желтой окраски семян, а - зеленой окраски, В - гладкая форма семени, b - морщинистая. Скр
Лекция №18. Сцепленное наследование
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распред
Хромосомное определение пола
Большинство животных являются раздельнополыми организмами. Пол можно рассматривать как совокупность признаков и структур, обеспечивающих способ воспроизводства потомства и передачу наследственной и
Наследование признаков, сцепленных с полом
Установлено, что в половых хромосомах находятся гены, отвечающие не только за развитие половых, но и за формирование неполовых признаков (свертываемость крови, цвет зубной эмали, чувствительность к
Лекция №20. Взаимодействие генов
Многочисленные опыты подтвердили правильность установленных Менделем закономерностей. Вместе с тем, появились факты, показывающие, что полученные Менделем числовые соотношения при расщеплении гибри
ВИДЫ ВЗАИМОДЕЙСТВИЯ АЛЛЕЛЬНЫХ ГЕНОВ
Различают полное доминирование, неполное доминирование, кодоминирование, аллельное исключение.
Аллельными генами называются гены, расположенные в идентичных локусах гомоло
Полное доминирование
Полное доминирование - это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот прис
Неполное доминирование
Так называется вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа
Кодоминирование
Кодоминирование - вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву, и
Комплементарность
Комплементарность - вид взаимодействия неаллельных генов, при котором признак формируется в результате суммарного сочетания продуктов их доминантных аллелей. Имеет место при наслед
Эпистаз
Эпистаз - вид взаимодействия неаллельных генов, при котором одна пара генов подавляет (не дает проявиться в фенотипе) другую пару генов.
Ген-подавитель называют эп
Полимерия
Это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерное действие генов может быть кумулятив
Лекция №21. Изменчивость
Изменчивость - способность живых организмов приобретать новые признаки и свойства. Благодаря изменчивости, организмы могут приспосабливаться к изменяющимся условиям среды обитания.
Мутации
Мутации - это стойкие внезапно возникшие изменения структуры наследственного материала на различных уровнях его организации, приводящие к изменению тех или иных признаков организма
Генные мутации
Генные мутации - изменения структуры генов. Поскольку ген представляет собой участок молекулы ДНК, то генная мутация представляет собой изменения в нуклеотидном составе этого участ
Хромосомные мутации
Это изменения структуры хромосом. Перестройки могут осуществляться как в пределах одной хромосомы - внутрихромосомные мутации (делеция, инверсия, дупликация, инсерция), так и между хромосомами - ме
Геномные мутации
Геномной мутацией называется изменение числа хромосом. Геномные мутации возникают в результате нарушения нормального хода митоза или мейоза.
Гаплоидия - у
Нерасхождение половых хромосом во время мейоза у матери
Нерасхождение половых хромосом во время мейоза у отца
Р
♀46, XX
×
♂46, XY
Типы гамет
Закон гомологических рядов наследственной изменчивости Н.И. Вавилова
«Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллел
Искусственное получение мутаций
В природе постоянно идет спонтанный мутагенез, но спонтанные мутации - достаточно редкое явление, например, у дрозофилы мутация белых глаз образуется с частотой 1:100 000 гамет.
Факторы, в
Модификационная изменчивость
Модификационная изменчивость - это изменения признаков организмов, не обусловленные изменениями генотипа и возникающие под влиянием факторов внешней среды. Среда обитания играет бо
Вариационная кривая
На основании вариационного ряда строится вариационная кривая - графическое отображение частоты встречаемости каждой варианты.
Среднее значение признака встречается чаще, а
Полное доминирование – это такой вид взаимодействие аллельных генов, при котором проявление одного из аллелей гена (А) не зависит от наличия в генотипе аллеля (А`) Гомозиготы АА не отличаются по фенотипу от гетерозигот АА`.
Явление полного доминирования у человека встречается редко, что связано со свойствами доминирование.
Свойства доминирования:
Относительность – проявление доминантного гена зависет от качество второго аллельного гена (встречается при множеством аллелизме). Пример: Выработка лактозы, усвоение молока.
Нестойкость – это проявление доминантного гена, может зависеть от генотипа в целом. Пример: аллопеция у мужчин.
Обратимость – в зависимости от внешних условий, ген проявляет себя либо как доминантный, либо как рецессивный. Пример: Окраска цветков примулы.
Неполное доминирование – фенотип гетерозигот (ВВ`) отлиается от гомозигот (ВВ) м (В`В`)промежуточным проявление признакак
Пример: Брахидактилия, Серповидно-клеточная анемия (СКА), Синдром Марфана. Кодоминирование – это такой вид взаимодействия аллельных генов, при котором в каждый из аллельных генов проявляет свое действие.
Пример: Четвертая группа крови по системе АВ0
Сверхдоминирование – это такой вид взаимодействия аллельных генов, при котором доминантный ген в гетерозиготном состояния проявляется сильнее, чем в гомо-.
Пример: Устойчивость к малярии при СКА, усиленный иммунитет при брахидактилии, повышенная работоспособность и одаренность при синдроме Марфана.
Аллельное исключение – это такой вид взаимодействия аллельных генов, при котором один ген подавляется в независимости от доминантности.
Пример: субклиническая форма гемофилии у женщин - носительнице.
7. Наследование групп крови по системам ab0, Rh, mn.В мембранах эритроцитов содержатся особые вещества, обладающие антигенными свойствами, - агглютиногены. С ними могут реагировать растворенные в плазме специфические антитела, относящиеся к фракции глобулинов, - агглютинины. При реакции антиген - антитело между несколькими эритроцитами образуются мостики, и они слипаются.
|
Агглютиногены на эритроцитах |
Агглютинины |
||||||
|
Группа крови |
|||||||
|
отсутствуют (0) |
отсутствуют (0) |
||||||
Около 85% населения Европы имеет в эритроцитах антиген Rh и образует группу Rh -положителъных индивидуумов. Остальные люди из европейской популяции лишены этого антигена и являются Rh -отрицательными. Синтез антигена Rh контролируется доминантным аллелем D и происходит у лиц с генотипами DD и Dd. Резус-отрицательные люди являются рецессивными гомозиготами (dd). При беременности Rh -отрицательной женщины (dd) Rh -положительным плодом (мужчина DD или Dd, плод Dd) при нарушении целостности плаценты в родах Rh -положительные эритроциты плода проникают в организм матери и иммунизируют его. При последующей беременности Rh -положительным плодом (Dd) анти- Rh -антитела проникают через плаценту в организм плода и разрушают его эритроциты. Развивается гемолитическая болезнь новорожденного. Ведущим ее симптомом является тяжелая анемия.
Антигены M и N были основаны в 1927г. Ландштейнером они могут присутствовать в организме одновременно (MN) или порознь (M и N), но никогда не могут одновременно отсутствовать. Они содержатся в эритроцитах, а также в тканях различных органов. По хим. Составу относятся к полисахаридам.
|
L m L m гр. крови M L n L n гр. крови N L m L n гр. крови MN |