40. الجنس وطي وغير المتجانسة
في سياق التطور ، شكلت غالبية الكائنات ثنائية المسكن آلية لتحديد الجنس تضمن تكوين عدد متساوٍ من الذكور والإناث ، وهو أمر ضروري للتكاثر الذاتي الطبيعي للأنواع. يمكن أن يحدث تحديد الجنس في مراحل مختلفة من التكاثر. هناك ثلاثة أنواع رئيسية من التحديد:
1) شاذ ،عندما يتم تحديد جنس الفرد في عملية تطور الجنين. يعتمد تحديد الجنس في هذه الحالة بشكل كبير على البيئة الخارجية ؛
2) جري،عندما يتم تحديد جنس كائن الابنة المستقبلية أثناء التولد الجيني في والدي الفرد ؛
3) سينغاميكحيث يتم تحديد جنس الابنة في وقت اندماج الأمشاج. هذا هو أكثر أنواع تحديد الجنس شيوعًا ، وهو ما يميز الحيوانات من جميع الأنواع.
مع أنواع العزم الزوجية والمتزامنة ، يعتمد الجنس على كروموسومات جنسية معينة.
في إناث الثدييات ، في المجموعة ثنائية الصبغيات من الكروموسومات ، يتم عزل زوج من الجونوسومات (الكروموسومات الجنسية) من نفس الشكل ، والكروموسومات XX المعينة. تحتوي الذكور في النمط النووي على كروموسومات X و Y. تحتوي إناث الطيور على كروموسومين جنسيين مختلفين (XY) وذكور (XX). خلال فترة الانقسام الاختزالي (الانقسام الاختزالي) ، تشكل الثدييات الأنثوية نوعًا واحدًا من الأمشاج مع الكروموسوم X ، لذلك يُطلق على الجنس الأنثوي هيموجاميتيك.يشكل الذكور نوعين من الأمشاج مع كروموسومات X و Y ، لذلك يسمى الجنس الذكري غير متجانسة.يعتمد تحديد جنس الثدييات على الحيوانات المنوية التي ستخصب البويضة. إذا تم تخصيب البويضة بواسطة حيوان منوي يحتوي على الكروموسوم X ، يتم وضع أنثى ؛ إذا كان الحيوان المنوي يحمل كروموسوم Y ، يتم وضع الذكر.
مخطط تكوين الأمشاج في الانقسام الاختزالي للذكور والإناث ، تحديد الجنس أثناء الإخصاب على النحو التالي:
هناك احتمال متساوٍ لوضع ذكر أو أنثى ، لأن عدد الحيوانات المنوية في الذكور (بيض الطيور) مع X- و Y- كروموسومات هو نفسه.
عادة ما يولد الذكور أكثر بقليل من الإناث ، ولكن في سياق التطور ، تم تطوير آلية للحفاظ على النسبة العددية بين الجنسين ، وبالتالي ، بحلول سن البلوغ والنضج الفسيولوجي ، تنخفض هذه النسبة بسبب ارتفاع معدل وفيات الأطفال. ذكور.
43. تجارب على تنظيم نسبة العامة
44. وراثة علامات الحدود. بول
وراثة الخصائص الجنسية المحدودة
وراثة الصفات التي تتحكم فيها الجينات المترجمة في الجسيمات الذاتية ، ولكن تتجلى بشكل حصري أو في الغالب في جنس واحد ، هي وراثة محدودة الجنس.التقسيم وفقًا لهذه الميزات يتوافق مع قواعد مندل. هناك أيضًا علامات يتحكم فيها الجنس. تحدث بسبب الجينات الموجودة في الكروموسومات الجنسية ، لكنها تظهر بشكل مختلف في كلا الجنسين. تشمل العلامات المرضية أو غير المرغوب فيها التي تحد من جنس الحيوانات في الحيوانات نقص تنسج الضرع والحلمات ، ونقص تنسج الغدد التناسلية (الخصيتين والمبيضين) ، الخصيتين ، التخلف في أجزاء مختلفة من الأعضاء التناسلية في العجول (لوحظ بشكل رئيسي في الحيوانات ذات اللون الأبيض) ، تضيق المستقيم والرحم في الأبقار ، عدم تنسج القنوات القطعية للذئب ، الحمل المطول عند الإناث ، التشوهات في الحيوانات المنوية عند الذكور ، إلخ. معظم هذه الحالات الشاذة تسبب ضعفًا جزئيًا أو كليًا في الوظيفة الإنجابية.
مشكلة تنظيم الجنس
تنشأ مشكلة تنظيم الجنس من الحاجة إلى زيادة الإنتاج الحيواني عن طريق الحصول في الغالب على أفراد من نفس النوع ، مما يعطي غلة أعلى من الحليب واللحوم والصوف والبيض ، إلخ. وهكذا ، في تربية الأبقار الحلوب ، تكون ولادة العجول مرغوب فيه أكثر ، وفي تربية الأبقار ، فكيف تنمو بسرعة. من الأفضل تربية الثيران والأبقار ذات القيمة العالية ، يُنصح بالحصول على ذكور من أجل تكاثر أسرع لأنماطهم الجينية. في تربية دواجن البيض ، يكون الحصول على الدجاج أكثر اقتصادا. فيما يتعلق بهذه الاحتياجات العملية ، لا يسعى الباحثون فقط إلى فهم آليات تحديد الجنس ، ولكن أيضًا دراسة إمكانيات التنظيم الاصطناعي للجنس. وتجدر الإشارة إلى أنه فيما يتعلق بالحيوانات الكبيرة مع نمو الأجنة داخل الرحم ، لم يتم حل هذه المشكلة بعد. يمكن تنظيم نسبة الجنس في الثدييات عن طريق تقسيم الحيوانات المنوية إلى جزأين: الأول - يحتوي على كروموسوم X في الحيوانات المنوية والثاني - يحتوي على كروموسوم Y. إن إخصاب إناث أحد هذه الكسور سينتج ذرية من نفس الجنس. أجريت تجارب على فصل الحيوانات المنوية إلى الأجزاء المحددة بواسطة الطرد المركزي والرحلان الكهربي والترسيب (الترسيب) باستخدام هيستيدين الأحماض الأمينية. أدى تلقيح إناث الأرانب ، على سبيل المثال ، بجزء أخف وزنا وأكثر قدرة على الحركة إلى التحول نحو الجنس الذكري. ومع ذلك ، لم يحدث تحول كامل في نسبة الجنس. يتم تطوير طريقة للتحديد الكمي للحمض النووي في الحيوانات المنوية عن طريق قياس شدة التألق النووي. النتائج التي تم الحصول عليها ، وفقًا لمؤلفي هذه الطريقة ، يمكن أن تصبح شرطًا أساسيًا للتقسيم الناجح للحيوانات المنوية في الثدييات إلى حاملات كروموسوم X أو Y.
التوالد العذري. هذا هو تطور الكائن الحي دون الإخصاب. يمكن الحصول على أفراد من نفس الجنس عن طريق تطوير أجنة من الأب (الذكورة)أو الأم (التوليد)الأمشاج. لذلك ، بتوجيه من B. L. Astaurov ، أجريت تجارب على الذكورة في دودة القز. تعرض بيض دودة القز غير المخصب لصدمة حرارية وتشعيع بالأشعة السينية ، مما أدى إلى تدمير نواتها دون الإضرار بالسيتوبلازم. ثم تم تلقيح هذه البويضات. تشكلت البيضة الملقحة من اندماج نوى اثنين من الحيوانات المنوية التي اخترقت البويضة ، وكان لدى الأفراد الذين نشأوا منها علامات على النوع الأب فقط.
في نسخة أخرى من التجارب ، تم تسخين البويضات التي لم يتم تخصيبها ولم تخضع بعد لتقسيم الاختزال ، وبالتالي إيقاف الانقسام الاختزالي والحفاظ على ثنائية الصبغيات في مجموعة الكروموسوم. من بين هذه البويضات ، بدون إخصاب (التوالد العذري) ، تطورت الإناث فقط ، ورثت خصائص الأم. يحدث نمو العذراء (التوالد العذري) في الظروف الطبيعية أو العفوية في الطيور. توصل I. V. Kudryavtsev و Z.A Oshchepkova و A.K.Golubev et al. ، على أساس التجارب التي أجريت على دجاج من سلالات مختلفة ، إلى استنتاج مفاده أن هناك شروطًا وراثية مسبقة للاختيار للحصول على أفراد قابلين للحياة - التوالد الفطري وإنشاء خطوط ذات قابلية عالية التوالد العذري. في هذه الحالة ، تتحول كل دجاجات الفقس إلى كوكيل. أدى الاختيار لزيادة وتيرة التوالد العذري ، الذي تم إجراؤه في سطرين من الديوك الرومية ، إلى النتائج التالية. زادت قدرة البيض على التوالد العذري من 1.1 إلى 18.6٪ في السطر الأول ومن 4 إلى 21.1٪ في السطر الثاني. لا تكمن أهمية التوالد العذري في أنه يسمح للشخص بالحصول على ذرية من نفس الجنس فحسب ، بل تكمن أيضًا في أن استخدام هذه الظاهرة يجعل من الممكن الحصول على أفراد متطابقين مع النمط الجيني لأحد الأشكال الأبوية.
يتم تمثيل الغالبية العظمى من الحيوانات من قبل أفراد من الجنسين - ذكور وإناث. في بعض أنواع النباتات ، لوحظ أيضًا أفراد من الجنسين (القنب ، طحلب الكبد spherocarpus ، المنوم ، إلخ). أصبحت الآلية التي تحدد جنس الفرد واضحة بعد الخلق نظرية الكروموسومالوراثة ، على الرغم من أن بعض الملاحظات حول أنماط وراثة الجنس قد تم إجراؤها قبل ذلك بكثير. من المعروف منذ فترة طويلة أنه في المتوسط ، في عدد الأنواع ، تكون نسبة الذكور والإناث هي نفسها ، أي تساوي 1: 1. هذه النسبة في G. (آه)والآخر متنحي متماثل الزيجوت (أأ).جعلت نسبة 1: 1 ، التي لوحظت عند التقسيم حسب الجنس ، من الممكن افتراض أن أحد الجنسين متغاير الزيجوت ، والآخر متماثل الزيجوت للعامل الذي يحدد جنس الكائن الحي. تم تأكيد هذا الافتراض في بداية القرن من خلال نتائج التجارب الجينية وتلقى تأكيدًا خلويًا بعد اكتشاف الكروموسومات الجنسية. عند دراسة تركيب كروموسومات الخلايا الجرثومية للحيوانات وخلايا أجسامها ، وجد أن الذكور والإناث يختلفون في مجموعة الكروموسومات. تمت دراسة الكروموسومات الجنسية بالتفصيل بواسطة W. Wilson و K. McKlang.
الكروموسومات الجنسية. التمثيل الحديثمن الملائم النظر في الآلية الصبغية لتحديد الجنس على كروموسومات ذبابة الفاكهة. على التين. 84 هو تمثيل تخطيطي لمجموعة الكروموسومات من ذبابة الفاكهة ، والتي تتكون من إجمالي ثمانية كروموسومات تشكل 4 أزواج. ثلاثة أزواج من الكروموسومات متجانسة مع بعضها البعض ولا تختلف شكليًا. تختلف كروموسومات الزوج الرابع اختلافًا كبيرًا. أحدهما يسمى كروموسوم X ، وهو على شكل قضيب ، والثاني (كروموسوم Y) له شكل خطاف مكون من ذراعي هذا الكروموسوم غير المتكافئ الحجم. في خلايا الإناث ، يتم تمثيل الزوج الرابع في ذبابة الفاكهة بواسطة اثنين من الكروموسومات X ، في الذكور
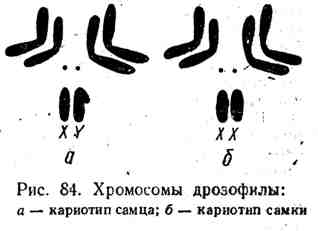
واحد X وكروموسوم Y واحد. يُطلق على زوج الكروموسومات التي يختلف بها الذكور والإناث الكروموسومات الجنسية أو الكروموسومات غير المتجانسة ، على عكس باقي الكروموسومات ، والتي تسمى autosomes (أحيانًا يتم اختصارها باسم A).
الجنس المثلي وغير المتجانسة. في إناث ذبابة الفاكهة ، والتي لها نفس الكروموسومات الجنسية (XX) ، جميع الخلايا الجرثومية الناتجة عن الانقسام الاختزالي لها نفس كروموسوم الجنس (X) ، بحيث تكون جميع الأمشاج متشابهة. يُطلق على الجنس الأنثوي من ذبابة الفاكهة ، الذي ينتج نوعًا واحدًا فقط من الأمشاج ، التماثل المتماثل. في المقابل ، ينتج ذكور ذبابة الفاكهة نوعين من الأمشاج ، أحدهما يتم تمثيل الكروموسوم الجنسي بالكروموسوم X ، والآخر بالكروموسوم Y. الجنس الذكري في ذبابة الفاكهة هو بالتالي غير متجانس.
اعتمادًا على ما إذا كانت الإناث تنتمي إلى الجنس المثلي أو غير المتجانسة ، يتم تقسيم جميع الحيوانات إلى مجموعتين. في الثدييات ، والديدان ، والقشريات ، ومعظم الحشرات (بما في ذلك ذبابة الفاكهة) ، ومعظم البرمائيات ، وبعض الأسماك ، يكون الذكر غير متجانس ، والأنثى متجانسة.
تشمل هذه المجموعة أيضًا البشر. تحتوي الخلايا الجسدية البشرية على 44 كروموسومًا وراثيًا ، بالإضافة إلى أن النساء لديهن كروموسومان X ، والرجال لديهم كروموسوم X واحد و كروموسوم Y واحد. يمكن تعيين صيغة الكروموسوم لجسم الإنسان الأنثوي 44A + XX، ذكر - 44 ألف + س ص. في الطيور والزواحف وبعض البرمائيات والأسماك وأجزاء من الحشرات (الفراشات وذباب الكاديس) يكون الجنس الأنثوي غير متجانس. في هذه الحالة ، يتم استخدام رموز أخرى لتعيين الكروموسومات الجنسية. عندما تكون الأنثى غير متجانسة ، فإن كروموسوماتها الجنسية تكون كذلك ZW, والكروموسومات الجنسية الذكرية ZZ. على سبيل المثال ، في الدجاج الذي يحتوي على 78 كروموسومًا في الخلايا الجسدية ، تكون صيغة الكروموسومات الذكرية -76A + ZZ، أنثى - 76 ألف + ZW.
في بعض الأنواع ، في عملية التطور ، كان هناك فقدان لأحد الكروموسومات الجنسية من الجنس غير المتجانسة ، ونتيجة لذلك في مجموعة الكروموسوميوجد كروموسوم جنسي واحد فقط. يشار إلى صيغة الكروموسومات الجنسية في هذه الحالة في الجنس غير المتجانسة على أنها XOأوZO.
آلية التقسيم حسب الجنس هي 1: 1. إن معرفة آلية تحديد الجنس بالكروموسومات تجعل من الممكن شرح أسباب ظهور الأفراد من الذكور والإناث بنسبة 1: 1. كما هو مذكور أعلاه ، فإن الجنس المتجانس يشكل أمشاج من نفس النوع ، كل منها يحمل نفس كروموسوم الجنس. في المقابل ، في الأمشاج من الجنس غير المتجانسة ، يتم تمثيل أحد الكروموسومات الجنسية أو الأخرى. على سبيل المثال ، في ذبابة الفاكهة ، تحتوي جميع الخلايا الجرثومية الأنثوية على كروموسوم X واحد ، ونصف الأمشاج الذكرية بها كروموسوم X ، والنصف الآخر يحتوي على كروموسوم Y.
إذا كانت البيضة (X) عن طريق الحيوانات المنوية مع كروموسوم X ، ثم يؤدي الجمع بين اثنين من الكروموسومات X في البيضة الملقحة إلى تكوين أنثى (XX). يؤدي إخصاب البويضة نفسها بالحيوان المنوي بالكروموسوم Y إلى ظهور الذكر (XY). نظرًا لحقيقة أن المشاركة في إخصاب حيوان منوي واحد أو آخر أمر محتمل بشكل متساوٍ ، فهناك احتمال متساوٍ لظهور الذكور والإناث. يمثل الجنس عدد كبيرحديثي الولادة في الحيوانات أنواع مختلفةتُظهر نسبة 1: 1 أو قريبة جدًا منها. لذلك ، من بين كل 100 مولود جديد ، يكون الذكور 51 في البشر ، و 56 في الكلاب ، و 49 في الدجاج ، و 50 في الفئران ، و 49 في الأغنام ، و 51 في الماشية ، و 52 في الخيول. ولادة. في المستقبل ، يمكن أن تتغير هذه النسب بشكل كبير بسبب عدم التكافؤ في بقاء الأفراد من الجنسين. لذلك في الشخص ، بحلول سن الخمسين ، تكون نسبة الرجال والنساء هي 85: 100 ، وبحلول عمر 85 - 50: 100.
يمكن تغيير مظهر جنس أو آخر ، الذي تحدده آلية الكروموسومات ، تجريبيًا في بعض الحالات ، كما هو موضح في الدجاج وديدان القز وما إلى ذلك. في النباتات ثنائية المسكن التي لها ذكور و نباتات أنثى، تمت أيضًا دراسة الكروموسومات الجنسية ؛ غير المتجانسة في معظم هذه النباتات هو الجنس الذكري.
معظم الحيوانات كائنات ثنائية المسكن. يمكن اعتبار الجنس على أنه مجموعة من الميزات والتراكيب التي توفر وسيلة لتكاثر النسل ونقل المعلومات الوراثية. غالبًا ما يتم تحديد الجنس في وقت الإخصاب ، أي أن النمط النووي للزيجوت يلعب الدور الرئيسي في تحديد الجنس. يحتوي النمط النووي لكل كائن حي على كروموسومات متماثلة لكلا الجنسين - الصبغيات ، والكروموسومات التي يختلف فيها الجنسين عن بعضهما البعض - الكروموسومات الجنسية. في البشر ، فإن الكروموسومات الجنسية "الأنثوية" هي نوعان من كروموسومات X. أثناء تكوين الأمشاج ، تتلقى كل بويضة أحد الكروموسومات X. يُطلق على الجنس الذي تتشكل فيه الأمشاج من نفس النوع ، وتحمل الكروموسوم X ، اسم homogametic. في البشر ، يكون الجنس الأنثوي متماثلًا. الكروموسومات الجنسية "الذكورية" في البشر هي كروموسوم X وكروموسوم Y. أثناء تكوين الأمشاج ، يتلقى نصف الحيوانات المنوية كروموسوم X ، والنصف الآخر - كروموسوم Y. يُطلق على الجنس الذي تتشكل فيه الأمشاج من أنواع مختلفة اسم غير متجانسة. في البشر ، يكون الجنس الذكري غير متجانس. إذا تم تكوين زيجوت يحمل اثنين من كروموسومات X ، فسيتم تكوين كائن حي منه ، إذا كان كروموسوم X وكروموسوم Y - ذكر.
الحيوانات لديها ما يلي أربعة أنواع من تحديد جنس الكروموسومات.
1. الجنس الأنثوي متماثل (XX) ، الذكر غير متجانس (XY) (الثدييات ، على وجه الخصوص ، البشر ، ذبابة الفاكهة).
المخطط الجيني لتحديد جنس الكروموسومات عند البشر:
المخطط الجيني لتحديد جنس الكروموسومات في ذبابة الفاكهة:
2. الجنس الأنثوي متماثل (XX) ، الذكر غير متجانس (X0) (Orthoptera).
المخطط الجيني لتحديد جنس الكروموسومات في الجراد الصحراوي:
3. الجنس الأنثوي غير متجانس (XY) ، الذكر متماثل (XX) (طيور ، زواحف).
المخطط الجيني لتحديد جنس الكروموسومات في الحمام:
Yandex.DirectAll الإعلانات
4. الجنس الأنثوي غير متجانس (X0) ، الذكر متماثل (XX) (بعض أنواع الحشرات).
علم الوراثة الجنسية
يتميز الجنس بمجموعة معقدة من السمات التي تحددها الجينات الموجودة على الكروموسومات. في خلايا جسم الإنسان ، تشكل الكروموسومات مجموعات ثنائية الصبغيات. في الأنواع ذات الأفراد ثنائي المسكن ، لا يكون مركب الكروموسومات للذكور والإناث هو نفسه ويختلف في زوج واحد من الكروموسومات (الكروموسومات الجنسية). كانت الكروموسومات المتطابقة لهذا الزوج تسمى كروموسوم X (x) ، غير مزدوج ، غائب في الجنس الآخر - Y (y) - كروموسوم ؛ الباقي ، الذي لا توجد اختلافات فيه ، هو جسمية (أ).
تحتوي خلايا المرأة على اثنين من الكروموسومات الجنسية المتطابقة ، والتي تم تحديدها XX ، ويتم تمثيلها في الرجال بواسطة كروموسومين X و Y غير متزاوجين. وهكذا ، فإن مجموعة الكروموسومات الخاصة بالرجل والمرأة تختلف في كروموسوم واحد فقط: مجموعة الكروموسوم للمرأة تحتوي على 44 جسمًا جسميًا + XX ، للرجال - 44 جسمًا جسميًا + XY.
أثناء انقسام ونضج الخلايا الجرثومية في البشر ، تتشكل الأمشاج مع عدد فردي من الكروموسومات: البيض ، كقاعدة عامة ، يحتوي على 22 + كروموسومات. وهكذا ، في النساء ، يتكون نوع واحد فقط من الأمشاج (الأمشاج مع كروموسوم X). في الذكور ، تحتوي الأمشاج على 22 + X أو 22 + كروموسومات Y ، ويتم تكوين نوعين من الأمشاج (مشيج كروموسوم X ومشيج كروموسوم Y). إذا دخل حيوان منوي به كروموسوم X البويضة أثناء الإخصاب ، يتم تكوين جنين أنثى ، ومع كروموسوم Y ذكر.
لذلك ، يعتمد تحديد جنس الشخص على وجود كروموسومات X أو Y في الخلايا الجنسية الذكرية - الحيوانات المنوية التي تخصب البويضة.
هناك أربعة أنواع رئيسية لتحديد جنس الكروموسومات:
1. الجنس الذكوري غير متجانس. 50٪ من الأمشاج تحمل كروموسوم X- ، 50٪ -Y ، على سبيل المثال ، البشر ، الثدييات ، Diptera ، الخنافس ، الحشرات (الشريحة 4).
2. الجنس الذكوري غير متجانس. 50٪ من الأمشاج تحمل X- ، 50٪ - لا تحتوي على كروموسوم جنسي ، على سبيل المثال ، الجنادب والكنغر (الشريحة 7).
3. الجنس الأنثوي غير متجانسة. 50٪ من الأمشاج تحمل X- ، 50٪ من الأمشاج تحمل الكروموسوم Y ، على سبيل المثال ، الطيور ، الزواحف ، البرمائيات الذيل ، دودة القز (الشريحة 7).
4. الجنس الأنثوي غير متجانسة. 50٪ من الأمشاج تحمل X- ، و 50٪ لا تحتوي على كروموسوم جنسي ، على سبيل المثال ، شامة.
وراثة الصفات التي تقع جيناتها على الكروموسومات الجنسية تسمى الوراثة ، المستعبدين على الأرض.
26. التركيب الجيني كنظام متكامل. تفاعل الجينات ، عمل متعدد للجينات.
التركيب الوراثي كنظام متكامل
خصائص الجينات.قائم على معرفةمع أمثلة وراثة الصفات في التهجينات أحادية وثنائية الهجين ، قد يحصل المرء على انطباع بأن النمط الجيني للكائن الحي يتكون من مجموع جينات فردية تعمل بشكل مستقل ، يحدد كل منها تطور سماته أو خصائصه فقط . إن فكرة وجود علاقة مباشرة لا لبس فيها بين الجين والسمة في أغلب الأحيان لا تتوافق مع الواقع. في الواقع ، هناك عدد هائل من سمات وخصائص الكائنات الحية التي يحددها زوجان أو أكثر من الجينات ، والعكس صحيح ، غالبًا ما يتحكم جين واحد في العديد من السمات. بالإضافة إلى ذلك ، يمكن تغيير عمل الجين بقرب الجينات الأخرى والظروف البيئية. وبالتالي ، ليس الجينات الفردية هي التي تعمل في مرحلة التطور ، ولكن النمط الوراثي بأكمله كنظام متكامل مع اتصالات معقدة وتفاعلات بين مكوناته. هذا النظام ديناميكي: ظهور أليلات أو جينات جديدة نتيجة للطفرات ، وتكوين كروموسومات جديدة وحتى جينومات جديدة تؤدي إلى تغيير ملحوظ في التركيب الجيني بمرور الوقت.
يمكن أن تتغير طبيعة مظهر عمل الجين في تكوين النمط الجيني كنظام في مواقف مختلفة وتحت تأثيرعوامل مختلفة. يمكن ملاحظة ذلك بسهولة إذا أخذنا في الاعتبار خصائص الجينات وخصائص مظاهرها في السمات:
الجين منفصل في عمله ، أي أنه معزول في نشاطه عن الجينات الأخرى.
الجين محدد في مظهره ، أي أنه مسؤول عن سمة أو خاصية محددة بدقة للكائن الحي.
يمكن للجين أن يعمل تدريجيًا ، أي زيادة درجة ظهور سمة مع زيادة عدد الأليلات السائدة (جرعة الجين).
يمكن لجين واحد أن يؤثر على التنمية علامات مختلفة- هذا عمل متعدد أو متعدد الاتجاهات للجين.
يمكن أن يكون للجينات المختلفة نفس التأثير على تطوير نفس الصفة (غالبًا سمات كمية) - هذه جينات متعددة ، أو جينات متعددة.
يمكن أن يتفاعل الجين مع الجينات الأخرى ، مما ينتج عنه سمات جديدة. يتم تنفيذ هذا التفاعل بشكل غير مباشر - من خلال منتجات ردود أفعالهم المركبة تحت سيطرتهم.
يمكن تعديل عمل الجين عن طريق تغيير موقعه في الكروموسوم (تأثير الموقع) أو بتأثير العوامل البيئية المختلفة.
تفاعلات الجينات الأليلية. تسمى الظاهرة عندما تكون عدة جينات (الأليلات) مسؤولة عن سمة واحدة تفاعل الجينات.إذا كانت هذه أليلات من نفس الجين ، فإن هذه التفاعلات تسمى أليليوفي حالة أليلات الجينات المختلفة - غير أليلي.
يتم تمييز الأنواع الرئيسية التالية من التفاعلات الأليلية: الهيمنة ، والسيطرة غير الكاملة ، والسيطرة المفرطة ، والسيطرة المشتركة.
هيمنة - نوع التفاعل بين أليلين لجين واحد ، عندما يستبعد أحدهما تمامًا مظهر عمل الآخر. هذه الظاهرة ممكنة في ظل الظروف التالية: 1) يوفر الأليل السائد في الحالة غير المتجانسة تخليق المنتجات الكافية لإظهار سمة من نفس الجودة كما هو الحال في حالة الزيجوت المتماثل السائد في الشكل الأبوي ؛ 2) الأليل المتنحي غير نشط تمامًا ، أو أن منتجات نشاطه لا تتفاعل مع نواتج نشاط الأليل السائد.
يمكن أن تكون أمثلة هذا التفاعل بين الجينات الأليلية هي هيمنة اللون الأرجواني تلوينزهور البازلاء على بيضاء ، وبذور ناعمة على التجاعيد ، والشعر الداكن على الضوء ، والعيون البنية على الأزرق في البشر ، وما إلى ذلك.
سيادة غير تامة، أو الطبيعة الوسيطة للميراث ،لوحظ في الحالة التي يختلف فيها النمط الظاهري للهجين (متغاير الزيجوت) عن النمط الظاهري لكل من الأبوين متجانسة الزيجوت ، أي أن التعبير عن السمة متوسط ، مع انحراف أكبر أو أقل تجاه أحد الوالدين أو الأب الآخر. آلية هذه الظاهرة هي أن الأليل المتنحي غير نشط ، ودرجة نشاط الأليل السائد غير كافية لتوفير المستوى المطلوب من مظاهر السمة السائدة.
مثال على الهيمنة غير المكتملة هو الميراث تلوينالزهور في نباتات جمال الليل (الشكل 3.5). كما يتضح من الرسم التخطيطي ، فإن النباتات متماثلة اللواقح إما حمراء (AA)إما بيضاء (آه)الزهور ، وغير المتجانسة (آه)- لون القرنفل. عند عبور نبات بأزهار حمراء ونبات به أزهار بيضاء في F 1 ، فإن جميع النباتات لها أزهار وردية ، أي الطبيعة الوسيطة للميراث.عند عبور الهجينة معالزهور الوردية في F 2 هناك مصادفة الانقسام حسب النمط الظاهري والنمط الجيني ، منذ الزيجوت المتماثل السائد (AA)يختلف عن متغاير الزيجوت (آه).لذلك ، في المثال قيد النظر مع نباتات جمال الليل ، تنقسم F 2 وفقًا للون الأزهار ، عادةً ما يكون ما يلي أحمرًا واحدًا (AA): 2لون القرنفل (آه): 1 أبيض (أأ).

أرز. 3. 5. وراثة لون الزهرة مع هيمنة غير كاملة في جمال ليلي.
ثبت أن الهيمنة غير المكتملة منتشرة على نطاق واسع. لوحظ في وراثة الشعر المجعد عند الإنسان ، ولون الماشية ، ولون ريش الدجاج ، والعديد من الخصائص المورفولوجية والفسيولوجية الأخرى في النباتات والحيوانات والبشر.
الهيمنة- مظهر أقوى من مظاهر السمة في فرد متغاير الزيجوت (آه)من أي متماثلة الزيجوت (AAو أأ).من المفترض أن هذه الظاهرة تكمن وراء التغاير (انظر الفقرة 3.7).
الترميز- مشاركة كلا الأليلين في تحديد الصفة في فرد متغاير الزيجوت. أحد الأمثلة المدروسة والمدروسة جيدًا للتشفير هو وراثة فصيلة الدم IV في البشر (المجموعة AB).
تحتوي كريات الدم الحمراء للأشخاص في هذه المجموعة على نوعين من المستضدات: المستضد أ(يحدده الجينوم / الموجود في أحد الكروموسومات) والمستضد في(يحدده الجين / أ ، المترجم في كروموسوم متماثل آخر). فقط في هذه الحالة يظهر كلا الأليلين تأثيرهما - 1 أ (الخامسمتماثل الزيجوت يتحكم في فصيلة الدم الثانية ، المجموعة أ) و أنا ب(في الحالة المتماثلة اللواقح يتحكم في فصيلة الدم الثالثة ، المجموعة ب). الأليلات 1 أو أنا بالعمل في الزيجوت متغاير الزيجوت كما لو كان بشكل مستقل عن بعضها البعض.
مثال الميراث مجموعاتيوضح الدم ومظهره allelism متعدد:يمكن تمثيل الجين بثلاثة أليلات مختلفة ، وهناك جينات تحتوي على عشرات الأليلات. يتم تسمية جميع الأليلات لجين واحد سلسلة من الأليلات المتعددة ،يمكن أن يكون لكل كائن حي ثنائي الصيغة الصبغية أي أليلين (وفقط). جميع المتغيرات المدرجة للتفاعلات الأليلية ممكنة بين هذه الأليلات.
ظاهرة الأليلية المتعددة شائعة في الطبيعة. من المعروف أن سلسلة واسعة من الأليلات المتعددة تحدد نوع التوافق أثناء الإخصاب في الفطريات ، والتلقيح في نباتات البذور ، وتحديد لون فراء الحيوانات ، إلخ.
التفاعلات الجينية غير الأليلية تم وصف التفاعلات الجينية غير الأليلية في العديد من النباتات والحيوانات. إنها تؤدي إلى ظهور نسل ثنائي الزيجوت من انقسام غير عادي وفقًا للنمط الظاهري: 9: 3: 4 ؛ 9: 6: 1 ؛ 13: 3 12: 3: 1 ؛ 15: 1 أي تعديلات على صيغة Mendelian العامة 9: 3: 3: 1. تُعرف حالات التفاعل بين اثنين أو ثلاثة أو أكثر من الجينات غير الأليلية. من بينها ، يمكن تمييز الأنواع الرئيسية التالية: التكامل ، والبلمرة.
مكملأو إضافي،يسمى هذا التفاعل بين الجينات السائدة غير الأليلية ، ونتيجة لذلك تظهر سمة غائبة في كلا الوالدين. على سبيل المثال ، عندما يتم عبور نوعين من البازلاء الحلوة مع الزهور البيضاء ، يتم إنتاج نسل من الزهور الأرجوانية. إذا قمنا بتعيين النمط الجيني لصنف واحد AAbb ،والآخر - aaBB ،الذي - التي
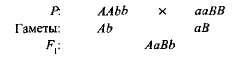
الجيل الأول هجين مع جينين مهيمنين (أو في)حصل على الأساس الكيميائي الحيوي لإنتاج أنثوسيانين الصباغ الأرجواني ، في حين لم يتم الحصول على جين واحد على حدة أ،لم يقدم أي من الجين B تركيب هذا الصباغ. تخليق الأنثوسيانين هو سلسلة معقدة من التفاعلات الكيميائية الحيوية المتسلسلة التي يتحكم فيها العديد من الجينات غير الأليلية ، وفقط في وجود اثنين على الأقل من الجينات المهيمنة (أ-ب-)يتطور اللون الأرجواني. في حالات أخرى (aaB-و أ-ب ب)زهور النبات بيضاء (تشير العلامة "-" في صيغة التركيب الوراثي إلى أن هذا المكان يمكن أن يشغله أليل سائد ومتنحي).
أثناء التلقيح الذاتي لنباتات البازلاء الحلوة F 1 الخامس F 2 لوحظ الانقسام إلى أشكال أزهار أرجوانية وبيضاء بنسبة قريبة من 9: 7. تم العثور على زهور أرجوانية في 9/1 6 نباتات بيضاء - في 7/16. تُظهر شبكة Punnett بشكل واضح سبب هذه الظاهرة (الشكل 3.6).
النخامة- هذا نوع من التفاعل الجيني حيث تقوم أليلات أحد الجينات بقمع التعبير عن الزوج الأليلي لجين آخر. الجيناتقمع عمل الجينات الأخرى يسمى مثبطات epistaticأو مثبطات.يسمى الجين المكبوت أقنومي.
وفقًا للتغير في عدد ونسبة النمط الظاهري وفئات chesk أثناء الانقسام الثنائي الهجين F 2 ضع في اعتبارك عدة أنواع من التفاعلات المعرفية: (أ> ب أو ب> أ)مع تقسيم 12: 3: 1 ؛ المتنحية (أ> بأو ب > أ) ،والتي يتم التعبير عنها في تقسيم 9: 3: 4 ، إلخ.
البلمرةيتجلى في حقيقة أن علامة واحدة تتشكل تحتها تأثيرعدة جينات لها نفس التعبير المظهرى. تسمى هذه الجينات البوليمر.في هذه الحالة ، يتم تبني مبدأ العمل الواضح للجينات على تطوير سمة. على سبيل المثال ، عند عبور نباتات كيس الراعي مع الفواكه المثلثة والبيضاوية (القرون) ، تتشكل النباتات ذات الثمار المثلثة في F 1. عندما يقومون بالتلقيح الذاتي في F 2 يوجد انقسام إلى نباتات ذات قرون مثلثة وبيضاوية بنسبة 15: 1. هذا لأن هناك نوعين من الجينات يعملان بشكل فريد. في هذه الحالات ، يتم تعيينهم على نفس النحو - أ 1 و أ 2 .

أرز. 3.6 . وراثة لون الزهرة في البازلاء الحلوة
ثم كل الأنماط الجينية (أ 1 ,-أ 2 ,-, أ 1 -أ 2 أ 2 ، أ 1 أ 1 أ 2 -) سيكون لها نفس النمط الظاهري - القرون المثلثة والنباتات فقط أ 1 أ 1 أ 2 أ 2 سوف تختلف - شكل القرون البيضاوية. هذا هو الحال بوليمر غير تراكمي.
يمكن أن تعمل الجينات البوليمرية أيضًا وفقًا للنوع بوليمر تراكمي.كلما كانت الجينات أكثر تشابهًا في النمط الجيني للكائن الحي ، كلما كان مظهر هذه السمة أقوى ، أي بزيادة جرعة الجين (أ 1 أ 2 أ 3 إلخ) عملها يتم تلخيصه أو تجميعه. على سبيل المثال ، تتناسب كثافة لون السويداء من حبوب القمح مع عدد الأليلات السائدة للجينات المختلفة في التهجين ثلاثي الهجين. كانت أكثر الحبوب الملونة أ 1 أ 1 أ 2 أ 2 أ 3 ،أ 3 أ الحبوب أ 1 أ 1 أ 2 أ 2 أ 3 أ 3 ليس لديها صبغة.
وفقًا لنوع البوليمر التراكمي ، يتم توريث العديد من السمات: إنتاج الحليب وإنتاج البيض والوزن والسمات الأخرى لحيوانات المزرعة ؛ العديد من المعلمات الهامة للقوة البدنية والقدرات الصحية والعقلية للشخص ؛ طول السنبلة في الحبوب ؛ محتوى السكر في جذور بنجر السكر أو الدهون فيه بذور زهرة عباد الشمسإلخ.
وبالتالي ، تشير العديد من الملاحظات إلى أن مظهر معظم السمات هو نتيجة لتأثير مجموعة من الجينات المتفاعلة والظروف البيئية على تكوين كل سمة محددة.
تفاعل الجينات
العلاقة بين الجينات والسمات معقدة للغاية. في الجسم ، لا يحدد جين واحد دائمًا سمة واحدة فقط ، وعلى العكس من ذلك ، يتم تحديد سمة واحدة بواسطة جين واحد فقط. في كثير من الأحيان ، يمكن لجين واحد أن يساهم في ظهور عدة سمات في وقت واحد ، والعكس صحيح. لا يمكن النظر إلى التركيب الجيني للكائن الحي على أنه مجموع بسيط من الجينات المستقلة ، كل منها يعمل بشكل مستقل عن الآخرين. المظاهر المظهرية لسمة معينة هي نتيجة تفاعل العديد من الجينات.
العمل المتعدد للجينات (تعدد الأشكال) - عمليات تأثير جين واحد على تكوين عدة سمات.
على سبيل المثال ، عند البشر ، يتسبب الجين الذي يحدد لون الشعر الأحمر في ظهور بشرة أفتح وظهور النمش.
في بعض الأحيان ، تؤثر الجينات التي تحدد السمات المورفولوجية على الوظائف الفسيولوجية ، وتقلل من القدرة على البقاء والخصوبة ، أو تتحول إلى قاتلة. لذلك ، فإن الجين الذي يسبب اللون الأزرق في المنك يقلل من خصوبته. تم تفصيل الجين السائد للون الرمادي في أغنام أستراخان في حالة متماثلة اللواقح ، لأن مثل هذه الحملان لديها معدة متخلفة وتموت عند التحول إلى تغذية العشب.
التفاعل التكميلي للجينات. يمكن أن تؤثر عدة جينات على تطور سمة واحدة. يسمى تفاعل العديد من الجينات غير الأليلية ، مما يؤدي إلى تطوير سمة واحدة ، التكميلية. على سبيل المثال ، يحتوي الدجاج على أربعة أشكال من القمة ، ويرتبط ظهور أي منها بتفاعل زوجين من الجينات غير الأليلية. المشط الوردي ناتج عن عمل الجين السائد لأليل واحد ، والمشط على شكل حبة البازلاء يرجع إلى الجين السائد للأليل الآخر. في الهجينة ، في وجود اثنين من الجينات غير الأليلية المهيمنة ، يتم تكوين مشط على شكل الجوز ، وفي حالة عدم وجود جميع الجينات السائدة ، أي في متماثل الزيجوت المتنحية لجينين غير أليليين ، يتم تكوين قمة بسيطة.
نتيجة تفاعل الجينات هو لون المعطف في الكلاب ، الفئران ، الخيول ، شكل اليقطين ، لون أزهار البازلاء الحلوة.
تعد البلمرة تفاعلًا بين الجينات غير الأليلية ، عندما تعتمد درجة تطور سمة ما على العدد الإجمالي للجينات السائدة. وفقًا لهذا المبدأ ، فإن لون حبوب الشوفان والقمح ولون البشرة عند الإنسان موروث. على سبيل المثال ، يمتلك السود 4 جينات سائدة في زوجين من الجينات غير الأليلية ، ولا يوجد لدى الأشخاص ذوي البشرة البيضاء ، جميع الجينات متنحية. مجموعات كمية مختلفةالمهيمن و الجينات المتنحيةيؤدي إلى تكوين الخلاسيين بكثافة مختلفة للون البشرة: من الداكن إلى الفاتح.
هناك مجموعتان رئيسيتان من التفاعل الجيني: التفاعل بين الجينات الأليلية والتفاعل بين الجينات غير الأليلية. ومع ذلك ، ينبغي أن يكون مفهوما أن هذا ليس كذلك التفاعل الجسديالجينات نفسها ، ولكن تفاعل المنتجات الأولية والثانوية التي ستحدد سمة أو أخرى. في السيتوبلازم ، يوجد تفاعل بين البروتينات - الإنزيمات ، التي يتم تحديد تركيبها بواسطة الجينات ، أو بين المواد التي تتشكل تحت تأثير هذه الإنزيمات.
الأنواع التالية من التفاعل ممكنة:
1) لتشكيل سمة معينة ، يكون تفاعل إنزيمين ضروريًا ، ويتم تحديد توليفهما بواسطة جينين غير أليلين ؛
2) إنزيم تم تصنيعه بمشاركة جين واحد يثبط أو يثبط نشاط الإنزيم الذي تم تكوينه بواسطة جين آخر غير أليلي ؛
3) إنزيمين يتم التحكم في تكوينهما بواسطة جينين غير متلازمين يؤثران على سمة واحدة أو عملية واحدة بحيث يؤدي عملهما المشترك إلى ظهور هذه السمة وتعزيزها.
تفاعل الجينات الأليلية
تسمى الجينات التي تحتل مواضع متطابقة (متجانسة) على الكروموسومات المتماثلة الأليلات. كل كائن حي لديه اثنين من الجينات الأليلية.
هذه الأشكال من التفاعل بين الجينات الأليلية معروفة: الهيمنة الكاملة ، والسيطرة غير الكاملة ، والسيطرة المشتركة والسيطرة المفرطة.
الشكل الرئيسي للتفاعل هو الهيمنة الكاملة ، والتي وصفها جي مندل لأول مرة. يكمن جوهرها في حقيقة أنه في كائن حي متغاير الزيجوت ، فإن مظهر أحد الأليلات يسيطر على مظهر الآخر. مع الهيمنة الكاملة ، لا يتطابق الانقسام حسب النمط الجيني 1: 2: 1 مع الانقسام حسب النمط الظاهري - 3: 1. في الممارسة الطبية ، من بين ألفي مرض وراثي أحادي الجين ، نصفهم تقريبًا لديهم مظاهر سائدة للجينات المرضية على الجينات الطبيعية. في الزيجوت المتغايرة ، يظهر الأليل المرضي في معظم الحالات كعلامات للمرض (النمط الظاهري السائد).
الهيمنة غير المكتملة هي شكل من أشكال التفاعل الذي لا يقوم فيه الجين السائد (A) في كائن حي متغاير الزيجوت (A) بقمع الجين المتنحي (أ) تمامًا ، ونتيجة لذلك تظهر وسيطة بين السمة الأبوية. هنا ، يتطابق الانقسام حسب النمط الجيني والنمط الظاهري ويكون 1: 2: 1
عندما سيطر عليها بشكل مشترك كائنات متغايرة الزيجوتيتسبب كل من الجينات الأليلية في تكوين منتج يعتمد عليه ، أي أن منتجات كلا الأليلين تكون كذلك. والمثال الكلاسيكي على مثل هذا المظهر هو نظام فصيلة الدم ، ولا سيما نظام ABO ، عندما تحمل كريات الدم الحمراء البشرية المستضدات السطحية التي يتحكم فيها كلا الأليلين. هذا الشكل من المظاهر يسمى الهيمنة المشتركة.
الهيمنة - متى الجين السائدفي حالة متغاير الزيجوت أكثر وضوحا مما كانت عليه في حالة متماثل الزيجوت. وهكذا ، فإن ذبابة الفاكهة ذات النمط الجيني AA لها متوسط عمر طبيعي متوقع ؛ أأ - التفاهة المطولة للحياة ؛ أأ - نتيجة قاتلة.
تعدد الولاء
كل كائن حي لديه اثنين فقط من الجينات الأليلية. في الوقت نفسه ، يمكن أن يكون عدد الأليلات في الطبيعة غالبًا أكثر من اثنين ، إذا كان بعض المواقع يمكن أن يكون في حالات مختلفة. في مثل هذه الحالات ، يتحدث المرء عن أليلات متعددة أو متعددة الأشكال.
يتم الإشارة إلى الأليلات المتعددة بحرف واحد بمؤشرات مختلفة ، على سبيل المثال: A ، A1 ، A3 ... يتم توطين الجينات الأليلية في نفس مناطق الكروموسومات المتجانسة. نظرًا لأن النمط النووي يحتوي دائمًا على اثنين من الكروموسومات المتجانسة ، حتى مع وجود أليلات متعددة ، يمكن أن يكون لكل كائن حي في وقت واحد فقط أليلين متطابقين أو مختلفين. يدخل واحد منهم فقط إلى الخلية الجرثومية (مع الاختلاف بين الكروموسومات المتجانسة). بالنسبة للأليلات المتعددة ، التأثير المميز لجميع الأليلات على نفس الصفة. الفرق بينهما هو فقط في درجة تطور السمة.
الميزة الثانية هي أن الخلايا الجسدية أو خلايا الكائنات ثنائية الصبغيات تحتوي على اثنين كحد أقصى من الأليلات المتعددة ، لأنها تقع في نفس موضع الكروموسوم.
ميزة أخرى متأصلة في الأليلات المتعددة. وفقًا لطبيعة الهيمنة ، يتم وضع الصفات المتغيرة الشكل في صف متسلسل: في كثير من الأحيان ، تهيمن سمة عادية غير متغيرة على الآخرين ، ويكون الجين الثاني من الصف متنحيًا بالنسبة إلى الأول ، ولكنه يهيمن على التالي ، إلخ. أحد الأمثلة على ظهور الأليلات المتعددة في البشر هو فصيلة الدم لنظام ABO.
تعد الأليلية المتعددة ذات أهمية بيولوجية وعملية كبيرة ، لأنها تعزز التباين التوليفي ، وخاصة التباين الوراثي.
تفاعل الجينات غير الآليل
تُعرف العديد من الحالات عندما يتم تحديد سمة أو خصائص بواسطة جينين أو أكثر من الجينات غير الأليلية التي تتفاعل مع بعضها البعض. على الرغم من أن التفاعل هنا مشروط أيضًا ، لأنه لا تتفاعل الجينات ، بل المنتجات التي تتحكم فيها. في هذه الحالة ، هناك انحراف عن أنماط الانقسام المندلية.
هناك أربعة أنواع رئيسية من التفاعل الجيني: التكامل ، والنقمة ، والبلمرة ، وتعديل الفعل (تعدد الأشكال).
التكامل هو نوع من تفاعل الجينات غير الأليلية ، عندما يكمل جين مهيمن عمل جين آخر غير أليلي سائد ، ويحددان معًا سمة جديدة غائبة عن الوالدين. علاوة على ذلك ، فإن السمة المقابلة تتطور فقط في وجود كل من الجينات غير الأليلية. على سبيل المثال ، يتم التحكم في لون طبقة الكبريت في الفئران بواسطة جينين (A و B). يحدد الجين A تخليق الصباغ ، ومع ذلك ، فإن كل من متجانسة الزيجوت (AA) ومتغايرة الزيجوت (Aa) هي ألبينو. يوفر جين B آخر تراكمات صبغية بشكل رئيسي في قاعدة الشعر ونهاياته. يؤدي عبور الزيجوت ثنائية الزيجوت (AaBb x AaBb) إلى تقسيم الهجينة بنسبة 9: 3: 4. يمكن أن تكون النسب العددية للتفاعلات التكميلية 9: 7 ؛ 9: 6: 1 (تقسيم مندلي المعدل).
مثال على التفاعل التكميلي للجينات في البشر يمكن أن يكون تخليق بروتين وقائي - الإنترفيرون. يرتبط تكوينه في الجسم بالتفاعل التكميلي لجينين غير أليليين يقعان على كروموسومات مختلفة.
التصلب هو تفاعل بين الجينات غير الأليلية حيث يقوم جين واحد بقمع عمل جين آخر غير أليلي. يمكن أن تسبب كل من الجينات السائدة والمتنحية (A> B ، a> B ، B> A ، B> A) الاضطهاد ، واعتمادًا على ذلك ، يتم تمييز النزف السائد والمتنحي. يسمى الجين الكابت بالمثبط أو الكابت. لا تحدد الجينات المانع بشكل عام تطور سمة معينة ، ولكنها تقوم فقط بقمع عمل جين آخر.
يسمى الجين الذي يتم قمع تأثيره باسم hypostatic. مع التفاعل المعرفي للجينات ، يكون الانقسام حسب النمط الظاهري في F2 هو 13: 3 ؛ 12: 3: 1 أو 9: 3: 4 ، إلخ. يتم تحديد لون ثمار اليقطين ولون الخيول من خلال هذا النوع من التفاعل.