Ein Gynoeceum ist eine Ansammlung von Megasporophyllen oder Fruchtblättern einer Blüte, die darin einen oder mehrere Stempel (Fruchtblätter) bilden.
Die blättrige Natur der Fruchtblätter wird bewiesen durch: 1) ihre Ähnlichkeit bei einer Reihe von Pflanzen (Hülsenfrüchte, einige Butterblumen usw.) in der Form mit einem gekräuselten Blatt; 2) anatomische Struktur- das Vorhandensein von Mesophyll und Stomata in der Epidermis; 3) die Wachstumsweise, die auch für Blätter charakteristisch ist; 4) das Fehlen einer Verschmelzung der Ränder der Fruchtblätter in den allerersten Entwicklungsstadien einiger polykarpöser Hülsenfrüchte und deren Verschmelzung erst in der Zukunft;
5) zahlreiche teratologische Fälle, bei denen die Fruchtblätter eine mehr oder weniger blattartige Form annehmen.
Im Stempel unterscheiden sie (Abb. 289) den bedeutendsten unteren, geschwollenen, hohlen Teil - den Eierstock, der im Inneren Megasporangien oder Eizellen enthält; Oberhalb des Eierstocks befindet sich normalerweise eine dünne zylindrische Säule, und darüber befindet sich eine Narbe sehr unterschiedlicher Form. Wenn der Eierstock ungleichmäßig wächst, kann die Säule seitlich oder an der Basis herausragen (bei Labiales, Borretsch, einigen Rosengewächsen). Bei einigen Pflanzen ist der Griffel nicht entwickelt, die Narbe befindet sich direkt am Fruchtknoten und wird als sitzend bezeichnet. Da die Eizellen im Eierstock eingeschlossen sind, können Staubpartikel nicht wie bei Gymnospermen direkt auf sie übertragen werden. Sie fallen auf andere Weise auf der Narbe (Bestäubung) und scheidet meist eine klebrige Flüssigkeit aus. Für einen größeren Bestäubungserfolg hebt die Säule die Narbe an.
In einfacheren Fällen wird der Stempel von einem Fruchtblatt gebildet (z. B. bei Motten), das konkav wird, sich zusammenrollt, seine Ränder zusammenlaufen und zusammenwachsen und an der Verschmelzungsstelle die sogenannte ventrale Naht bilden (Abb. 290). . Auf der der ventralen Naht gegenüberliegenden Seite befindet sich die Mittelader des Fruchtblattes, die fälschlicherweise als dorsale Naht bezeichnet wird. Befinden sich in der Blüte mehrere solcher Einzelstempel, so nennt man das gesamte Gynoeceum apokarpös (in
H- Eierstock; st- Spalte; R- Schnauze.

1 - Eizellen befinden sich fast entlang der gesamten Innenfläche des Eierstocks; 2 - Die Samenanlagen befinden sich an den Rändern des Fruchtblatts, der Eierstock ist entlang der Bauchnaht künstlich geöffnet.

1 - apokarpös; 2-4 - känokarpös in unterschiedlichem Verschmelzungsgrad der Fruchtblätter; H- Eierstock; st- Spalte; R- Schnauze.
Ranunculaceae, viele Rosaceae usw.). Wenn die Blüte mehrere Fruchtblätter enthält, wachsen sie in den meisten Fällen zusammen und bilden einen Stempel. Ein Gynoeceum, das aus mehreren verwachsenen Fruchtblättern besteht, wird stellifoliös oder synkarpös, besser gesagt coenokarpös genannt. Die Verschmelzung der Fruchtblätter erfolgt meist von unten nach oben. Es kann sich (Abb. 291) nur auf die Eierstöcke (Nelken, Flachs, Rhabarber usw.), auf die Eierstöcke und Säulen in ihrem unteren Teil oder auf die Narben selbst (Safran, Iris, Compositae, Labiales usw.) ausbreiten , schließlich an Eierstöcken, Säulen und Narben (Primeln). Da der Eierstock am meisten ist wichtiger Teil Stempel, dann wird die Anzahl der Stempel in einer Blüte durch die Anzahl der Eierstöcke und nicht durch die Anzahl der Säulen bestimmt. Anhand unverwachsener Griffel, Narben oder Narbenlappen kann man die Anzahl der Fruchtblätter, die den Stempel bilden, beurteilen (aber nicht immer genau). Im Falle ihrer vollständigen Verschmelzung kann die Anzahl der Fruchtblätter anhand der Anzahl ihrer Mitteladern in der Fruchtknotenwand beurteilt werden. Bei sehr wenigen Pflanzen (Gushs, Kutrs) wachsen die oberen Teile der Fruchtblätter zusammen – die Säulen und Narben oder nur die Narben und die Fruchtknoten bleiben frei. In diesen Fällen wird auch davon ausgegangen, dass sich in der Blüte nur ein Stempel befindet.
Eierstock. Plazenta. Eierstockstellen, an denen die Eizellen befestigt sind und über die sie versorgt werden Nährstoffe werden Plazenta oder Plazenta genannt. Samenanlagen (Megasporangien) im Fruchtknoten können wie Sporangien an jeder Stelle des Fruchtblatts entstehen. Aber nur bei sehr wenigen Pflanzen, hauptsächlich aus den primitivsten Familien – Seerosen, Susaks und anderen – befinden sie sich auf einem erheblichen Teil der Innenfläche der Fruchtblätter (laminal-laterale Plazenta, Abb. 292, 10 ). Normalerweise entwickeln sich die Samenanlagen auf der Innenseite der Fruchtblätter nur in der Nähe ihrer äußersten Ränder und bilden zwei (bei Hülsenfrüchten, Lilien) oder mehr Längsreihen entlang der Bauchnaht oder befinden sich dort ohne jegliche Ordnung.
Im Verlauf der Evolution des Gynäkiums wandelte es sich in verschiedenen Evolutionsreihen von apokarpös zu känokarpös um, d. h. die Fruchtblätter wuchsen zusammen und bildeten einen Stempel. Gleichzeitig biegen sich bei sehr vielen Pflanzen die Ränder der Fruchtblätter nach innen, verwachsen dort mit ihren Seitenflächen und bilden einen Fruchtknoten, der in Kammern, sogenannte Nester, unterteilt ist. verschmolzen Seitenflächen Fruchtblätter
erreichen die Mitte und bilden Trennwände im Eierstock, die ihn in eine Anzahl von Nestern unterteilen, die der Anzahl der Fruchtblätter entspricht; Eierstöcke sind 2-, 3-, 4-, 5-, 6- und mehrzellig (z. B. bei Lilien, Huftieren, Geranien, Glockenblumen1 usw.).
Ein solches Gynoeceum wird im engeren Sinne synkarpös genannt. Plazenten mit Eizellen in solchen Eierstöcken befinden sich in den inneren Ecken der Nester des Eierstocks, entlang der Ränder der hier umwickelten Fruchtblätter und werden als zentral oder besser bezeichnet bezeichnet zentrale Ecke Plazenten (in Lilien, Campanulas usw., Abb. 292, 3-4 ); dabei
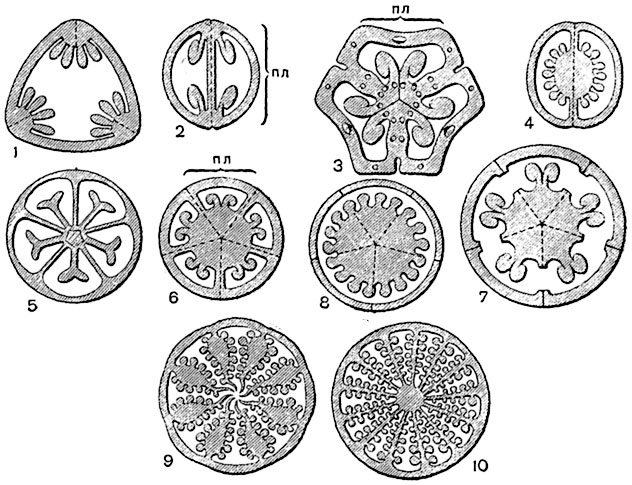
1-2 - Plazenta parietal ( 2 - mit einer falschen Partition); 3-5 - Plazenta zentraleckig; 6 - anfängliche und 7 - das letzte Entwicklungsstadium der falschaxialen Plazenta; 8 - falschaxialer Samenträger; 9 - 10 - Plazenta befinden sich entlang der Trennwände in nicht ganz ( 9 ) und ganz ( 10 ) mehrzelliger Eierstock; pl- Fruchtblatt.
In einigen Fällen (Nachtschatten, Heidekraut usw.) wachsen die Plazenten stark und ragen weit in die Höhle der Eierstocknester hinein. Bei vielen Pflanzen (Labiaceae, Borretsch, Flachs usw.) entwickeln sich im Eierstock zusätzlich zu den „echten“ Trennwänden, die durch die umwickelten Ränder der Fruchtblätter gebildet werden, auch falsche Trennwände aus Auswüchsen der Innenwand des Fruchtknotens; Daher ist es nicht immer möglich, anhand der Anzahl der Nester im Eierstock die Anzahl der Fruchtblätter zu beurteilen, die ihn gebildet haben.
Im Prozess der weiteren Entwicklung des synkarpösen Gynoeceums in verschiedenen Evolutionsreihen Angiospermen es kam zu einer Umwandlung eines zwei-, mehrzelligen Eierstocks in einen einzelligen: Die nach innen gewickelten und verwachsenen Fruchtblätter trennten sich, richteten sich auf, blieben aber mit ihren Rändern miteinander verwachsen; Es wurde ein einzelliger Fruchtknoten erhalten, der aus mehreren Fruchtblättern besteht, die mit ihren Rändern nur entlang der Peripherie des Fruchtknotens miteinander verwachsen (z. B. bei Veilchen, Johannisbeeren, Stachelbeeren, Weiden, Enzianen, Sommerwurz, fast allen Orchideen, usw.). Diese Art von känokarpösem Gynoeceum wird parakarpös genannt. Plazenten befinden sich in diesem Fall entlang der Innenwände des Eierstocks (wo die Ränder der Fruchtblätter zusammengewachsen sind) und werden als Parietal, Wand oder häufiger als Parietal bezeichnet. Bei einer Reihe von Pflanzen mit parietalen Plazenten ragen diese stark in die Eierstockhöhle hinein und erwecken den Eindruck falscher Trennwände (Kürbis, Mohn usw., Abb. 292, 9 ). Und im parakarpösen Gynoeceum im Eierstock können sich aus Auswüchsen der Innenwand des Eierstocks falsche Trennwände bilden, wie zum Beispiel bei Kreuzblütlern (Abb. 292, 2 ), wobei die Scheitelplazenta die Verschmelzungsstellen der Ränder der Fruchtblätter anzeigt. Durch die Reduktion verringerte sich die Anzahl der Eizellen im Eierstock des parakarpösen Gynoeceums in einigen Familien auf eins (Asteraceae, Gräser usw.).
Parietale Plazenten kommen auch in einem einzelligen Eierstock mit vielen Eizellen vor, die von einem Fruchtblatt gebildet werden, wie zum Beispiel bei Hülsenfrüchten, Pfingstrosen, Winterfrüchten usw.
Aus dem synkarpösen Gynoeceum ist in manchen Evolutionsreihen auch das sogenannte lysicarische Gynoeceum hervorgegangen. Hier wuchsen die Ränder der Fruchtblätter, die sich in sehr frühen Stadien der Ontogenese nach innen drehten und verschmolzen, später nicht und bildeten keine Trennwände im Eierstock, der einzellig sein wird. Von der Unterseite des Eierstocks erhebt sich die Plazenta in Form einer Säule, als wäre sie ein Gefäß. In Wirklichkeit wird auch hier die Plazenta durch die miteinander verwachsenen Ränder der Fruchtblätter gebildet (Abb. 292, 6-8 ). Es ist möglich, dass bei manchen Pflanzen das Gefäß in der Mitte des Fruchtknotens zwischen den Enden der Fruchtblätter wächst, es aber trotzdem an der Peripherie mit Fruchtblattgewebe bedeckt ist. Eine solche Plazenta wird axial, falschaxial, freizentral, säulenförmig 10 genannt. Es ist typisch für Primeln, einige Nelken, Blei, Buchweizen (bei letzteren ist die Anzahl der Eizellen auf eins reduziert) und einige andere.
Die Anzahl der Eizellen im Eierstock verschiedener Pflanzen reicht von einer bis zu mehreren Tausend.
Das primitivste wird ein apokarpöses Gynoeceum mit vielen Eizellen in jedem einzelligen Fruchtknoten sein, die aus einem einzelnen Fruchtblatt bestehen. Im Laufe der Evolution verringerte sich die Anzahl seiner Fruchtblätter auf 3-2 und schließlich auf eins (bei Hülsenfrüchten, Rittersporn, Pflaumen, Kirschen usw.), und auch die Anzahl der Samenanlagen im Fruchtknoten verringerte sich schließlich auf eine (Butterblumen, Anemonen, die viele Stempel in einer Blüte haben, Pflaume, Kirsche, die einen Stempel pro Blüte haben usw.). Andererseits entwickelte sich im Laufe der Evolution aus dem apokarpösen Gynoeceum ein synkarpöses (im engeren Sinne) und aus diesem parakarpöses und lysikarpöses Gynäkium. Und bei diesen känokarpösen (wachstumsblättrigen) Gynoeceien verlief die Evolution hauptsächlich auf dem Weg einer allmählichen Verringerung der Anzahl der Fruchtblätter (auf zwei) und der Anzahl der Eizellen (auf eins).
Es ist jedoch wahrscheinlich, dass beispielsweise bei Orchideen eine kolossale Anzahl von Eizellen im Fruchtknoten aus einer Größenabnahme und einer Zunahme der Anzahl der ursprünglich nicht so vielen Eizellen ihrer Vorfahren resultierte.
Der sowjetische Botaniker A. L. Takhtadzhyan hat kürzlich die Entwicklung von Gynäkium- und Plazentationstypen untersucht.
Abhängig von der Position des Fruchtknotens im Verhältnis zum Rest der Blüte kann er oben, unten und halb unten liegen. Der obere Eierstock wird frei auf einem konvexen, flachen oder konkaven Gefäß sitzend genannt; seine Wand besteht nur aus Fruchtblättern. Befindet sich ein solcher Eierstock an der Oberseite des Gefäßes und sind die übrigen Blütenglieder darunter befestigt, dann wird die Blüte Pistillat genannt (Abb. 293, 1 ). Eine andere Art von oberem Eierstock liegt vor, wenn er am Boden einer konkaven Kannenformation sitzt, ohne mit dieser zu verwachsen (z. B. bei Hagebutten, Kirschen, Pflaumen, Pfirsichen, Aprikosen, Manschetten usw.). Dieser Krug besteht aus einem einzelnen konkaven Gefäß oder, wie bei der Wildrose, aus einem unteren Teil aus einem konkaven Gefäß und dem oberen Teil aus den miteinander verwachsenen Basen der Blütenblätter und Staubblätter. Oft wird ein solcher Eierstock nicht als oberer, sondern als mittlerer Eierstock bezeichnet. Die übrigen Blütenglieder, die am oberen Rand des Kruges befestigt sind, befinden sich in diesem Fall auf Höhe der Oberseite des Fruchtknotens (aber nicht darauf), und die Blüte befindet sich in der Nähe des Stempels (Abb. 293, 2, 3 ).

1 - oberer Fruchtknoten, pistillierte Blüte; 2-3 - oberer (oder mittlerer) Eierstock, Blüte in der Nähe des Stempels; 4 - der untere Fruchtknoten, der aus dem Gefäß und den Fruchtblättern besteht; die Blüte ist supragillär; 5 - der untere Fruchtknoten besteht aus Fruchtblättern, die mit den unteren Teilen der Kelchblätter, Blütenblätter und Staubblätter verwachsen sind; die Blüte ist supragillär; 6 - halbunterer Fruchtknoten, halbadaptive Blüte. Feige. 1-5 skizzenhaft.
Der untere Eierstock wird als einer bezeichnet, an dessen Spitze sich eine Blütenhülle befindet, und er geht genau von der Spitze des Eierstocks selbst ab und nicht von der Spitze des Gefäßes, das nicht mit ihm verschmolzen ist, wie im vorherigen Fall. Die Blüte hier wird supraspinal genannt.
Der Ursprung eines solchen Eierstocks ist unterschiedlich. In einigen Fällen werden die Wände des unteren Fruchtknotens von Fruchtblättern und einem damit verwachsenen konkaven Kelchgefäß gebildet, z. B. bei Laurentaceae (Abb. 293, 4 ). In den allermeisten Fällen ist das Gefäß nach neueren Studien nicht an der Bildung des unteren Eierstocks beteiligt: Es wird nur von den unteren Teilen der Blütenhülle, den Staubblättern und den Fruchtblättern gebildet (Abb. 293, 5 ). Sogar bei Äpfeln, Birnen und Quitten stellt sich heraus, dass der untere Fruchtknoten, der bis vor Kurzem durch die Verschmelzung eines konkaven Gefäßes mit Fruchtblättern gebildet wurde, neuen Studien zufolge nicht durch das Gefäß, sondern durch die Basen der Kelchblätter gebildet wird , Blütenblätter, Staubblätter, miteinander und mit Fruchtblättern verwachsen.
Dieser oder jener Ursprung des unteren Eierstocks wird durch eine gründliche vergleichende Untersuchung des Durchgangs von Gefäßbündeln in ihm in einer Reihe eng verwandter Gattungen geklärt.
Bei einem semi-inferioren Eierstock ist er nur in seinem unteren Teil mit dem Gefäß oder mit der Blütenhülle und den Staubgefäßen verwachsen, während sein oberer Teil frei ist (Abb. 293, 6 ); Die Blütenhülle geht in diesem Fall von der Mitte des Eierstocks ab und wird als Blüte bezeichnet halbklebend(in Holunder, Geißblatt, Steinbrech).
Obere und untere Eierstöcke sind sehr häufig, halbuntere kommen bei wenigen Pflanzen vor. Dieser oder jener Eierstocktyp ist erblich konstant und für die Taxonomie von großer Bedeutung.
Zwischen all diesen Eierstocktypen gibt es Übergänge. Unter dem Gesichtspunkt der vergleichenden Morphologie können die unteren und halbunteren Eierstöcke nicht als vollständig homolog zu den oberen angesehen werden, da an ihrer Bildung neben den Fruchtblättern auch das Gefäß oder die Tepalen und Staubblätter beteiligt sind.
Phylogenetisch ist der obere Eierstock älter als der untere. Der untere Eierstock kann, wie bereits angedeutet, einen anderen Ursprung haben. Im Laufe der Evolution der Angiospermen ist der untere Eierstock in seinen verschiedenen phylogenetischen Reihen immer wieder und unabhängig voneinander entstanden.
Samenanlage. Die Eizelle oder Megasporangium von Angiospermen besteht im Allgemeinen aus den gleichen Teilen wie die von Gymnospermen: dem Samenstiel, der sie an der Samenkapsel befestigt, dem Nucellus (oder Zellkern) und einer (in fast allen interpetalen) oder zwei Hüllen, oder Hüllen (bei Monokotyledonen und fast allen einzelnen Blütenblättern); Letztere schließen sich oben nicht und hinterlassen ein Loch – den Polleneingang (Mikropyle). Nur bei sehr wenigen (Santalaceae, Balanophora, Limbata usw.) sind die Samenanlagen kahl, ohne Hülle.
Der Kern bei zweihäusigen und den meisten einkeimblättrigen Arten ist gut entwickelt, besteht meist aus vielen Zellreihen und bleibt lange erhalten ( krass Eizellen). Bei Spaltungen ist es meist schlecht entwickelt, besteht aus wenigen Zellen, und wenn sich darin der Embryosack entwickelt, wird er von diesem verdrängt, ernährt sich von ihm und wird fast spurlos zerstört ( tenuinucelates Eizellen). Der erste Nucellustyp muss als primitiver angesehen werden und seine Reduzierung erfolgte als Ergebnis der weiteren Evolution.
Die Stelle, an der die Eizelle am Samenstiel befestigt ist, wird Narbe genannt (sie ist auf den Samen deutlich sichtbar), und die Basis des Nucellus, von der sich die Haut erstreckt, ist Chalatsa. Die Eizelle wird in Form eines Tuberkels aus einem Dermatogen und einem Periblem gelegt; später bilden sich aus seiner Basis - chalatsy - Integumente, in den meisten Fällen ist es basipetal, d. h. das innere ist früher als das äußere.
Die Samenanlage kann mit dem Sporangiumstiel, der Nucellus mit der Megasporangiumwand homologiert werden. Manche Decken (Integumente) ähneln den Decken (Induums) von Farnen. Viele halten sie für ein Neoplasma, da sie aus der Eizelle selbst gebildet werden und das Hochblatt von Farnen einen Sporophyllauswuchs aufweist.
Es ist auch möglich, wie bereits in der Diskussion der Gymnospermen (S. 315) dargelegt wurde, dass es sich bei den Integumenten der Samenanlage um steril gewordene und reduzierte Reste von Megasporangien handelt, die mit dem zentralen Megasporangium verschmolzen waren. aus dem der Nucellus entstand, zu einem Synangium. Mit diesem Verständnis des Ursprungs des Integuments muss streng genommen nur der Nucellus als Homolog des Megasporangiums betrachtet werden, und die gesamte Eizelle wird homolog zum Sorus oder, genauer gesagt, zum Synangium der Megasporangia sein von denen nur eine eine Megaspore enthält und die anderen reduziert und in eine Hülle (oder zwei Hüllen) umgewandelt werden. ).
Überblick über den Generationswechsel und den Wechsel der Kernphasen in verschiedenen Pflanzengruppen

Reduktionsabteilung

Düngung
Früher galt die Samenanlage als modifizierte Knospe, daher der Name; Nucellus galt als Stängel und Integument als Blätter. Beweise für den mittlerweile akzeptierten Ursprung der Samenanlagen als Auswüchse an den Rändern oder auf der Oberfläche der Fruchtblätter sind: 1) die Umwandlung der Lappen des Fruchtblatts in Samenanlagen in den Cycas (siehe S. 314); 2) die Lage der Samenanlagen bei vielen Pflanzen auf der Parietalplazenta, d. h. entlang der Ränder der Fruchtblätter; 3) basipetale Entwicklung der Eihüllen, während sich die Blätter am Stängel akropetal entwickeln; 4) Übergangsformationen in grünlichen Blüten (S. 325) zwischen Lappen, Zähnchen, Auswüchsen von Fruchtblättern und Samenanlagen; 5) manchmal beobachtete Fälle von missgebildeten Staubgefäßen, bei denen das Staubblatt (Pollen) teils Pollennester, teils Eizellen bildet; 6) die Möglichkeit der Homologation der Eizelle mit Megasporangium heterosporiger Farne usw. Daher ist es möglicherweise richtiger, die Eizelle, die in uns keine Wurzeln geschlagen hat, zu benennen - Eizelle oder Eizelle.

Reis. 294. Die wichtigsten Arten von Eizellen und ihre Entwicklung (Schema).
Wie ist die Struktur einer Blume?
Blume - Orgel Samenvermehrung. In der Blüte werden folgende Teile unterschieden (von der Peripherie zur Mitte):
1) Blütenhülle;
1) Staubblätter;
3) Stößel (oder Stößel).
Wenn die Blütenhülle aus einem Kelch und einer Krone besteht, wird sie beispielsweise bei einem Apfelbaum, einer Erbse, einer Glockenblume oder einer Kartoffel als doppelt bezeichnet. Manchmal ist die Blütenhülle nicht in Kelch und Krone unterteilt. Eine solche Blütenhülle wird beispielsweise bei einer Tulpe, einem Maiglöckchen, einer Zwiebel, einer Eiche oder einer Birke als einfach bezeichnet. Einige Pflanzen haben keine Blütenhülle (die Blüten sind „nackt“): Weide, Pappel.
Besteht der Kelch aus einzelnen Kelchblättern, spricht man von Einzelblättrigen (Wilder Rettich, Erdbeere). Wenn die Kelchblätter miteinander verschmelzen, ist der Kelch sympathisch (Erbse, Nelke). Außerdem kann die Blütenkrone aus einzelnen Blütenblättern (Apfel, Wildrose, Butterblume) und sympetal (Glocke, Tabak, Flieder) bestehen.
Wenn die Blüten mehrere Symmetrieebenen haben, werden solche Blüten als regelmäßig bezeichnet (Lilie, Nelke, Gurke). Manchmal haben Blumen nur eine Symmetrieebene. Solche Blüten nennt man unregelmäßig (Salbei, Erbse, Löwenmäulchen).
Das Staubblatt besteht aus einem Staubbeutel an einem Staubfaden. Pollen werden im Staubbeutel produziert.
Der Stempel besteht aus Narbe, Griffel und Fruchtknoten. Im Eierstock bilden sich Eizellen, aus denen nach Bestäubung und Befruchtung Samen entstehen. Der Eierstock ist oben und unten. Der obere Fruchtknoten liegt über dem Rest der Blüte. Es ist frei, es ist leicht von der Blume zu unterscheiden. Blüten mit oberem Eierstock - Erbsen, Kartoffeln, Radieschen, Nelken. Der untere Fruchtknoten befindet sich unter der übrigen Blüte und verschmilzt mit dieser. Es kann nicht isoliert werden, ohne die Blüte zu zerstören. Der untere Eierstock befindet sich bei Sonnenblume, Gurke und Glockenblume.
Alle Teile der Blüte befinden sich am Gefäß – dem axialen Teil der Blüte. Unter der Blüte sind oft Blütenstiele deutlich sichtbar. Fehlen die Stiele, nennt man die Blüten sitzend (Wegerich).
Auf dieser Seite wurde gesucht nach:
- Wie ist die Struktur einer Blume?
- Aufsatz über die Struktur einer Blume
- Wie ist die Struktur einer Blume?
- Wie ist der Eierstock einer Blume aufgebaut?
- Aus welchen Teilen besteht eine Blume?
Aufgrund ihrer Zerbrechlichkeit (von mehreren Stunden bis zu mehreren Tagen und nur bei einigen tropischen Orchideen bis zu 80 Tagen) verändern sich Pflanzenblüten im Vergleich zu anderen Organen am wenigsten unter dem Einfluss äußerer Bedingungen. Daher haben systematische Botaniker sowohl in der Vergangenheit als auch heute ihre Klassifizierungssysteme für Angiospermen hauptsächlich auf der Grundlage der Struktur von Blüten erstellt und bauen diese weiter auf. Die Ähnlichkeit oder Nähe der Struktur von Blüten war so weit verbreitet, dass auf dieser Grundlage Arten nicht nur in Gattungen, sondern auch in Familien eingeteilt wurden, von denen viele sehr reichhaltig sind, zum Beispiel die Orchideenfamilie, die mehr zählt Über 20.000 Arten umfasst die Familie der Korbblütler, außerdem mehr als 20.000 Arten, die Familie der Nachtfalter usw. Einige dieser artenreichen Familien umfassen große Nummer wichtig medizinische Pflanzen Daher ist es angebracht, sich mit der typischsten Struktur ihrer Blüten und Früchte vertraut zu machen.
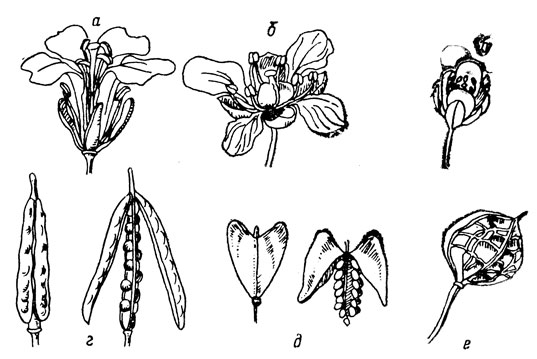
Familie der Kreuzblütler (Cruciferae). Eine Blüte bei Pflanzen dieser Familie hat den folgenden typischen Aufbau (Abb. 8): ein Kelch, bestehend aus 4 unverbundenen Kelchblättern, eine Blütenkrone, ebenfalls bestehend aus 4 Blütenblättern, unverbunden, unterschiedlich gefärbt, kreuzweise angeordnet, 6 Staubblätter in zwei Kreisen angeordnet – zwei kürzere im äußeren und vier längere im inneren Kreis (Vierstärke oder Tetradynamus), ein aus zwei Fruchtblättern bestehender Stempel mit einem oberen Fruchtknoten, eine trockene Frucht, die sich normalerweise entlang zweier Nähte öffnet, genannt a Schote (Siliqua) – in Senf, Kohl, Levkoy oder eine Schote (Silicula) im Hirtenbeutel, Getreide, Rote Bete.
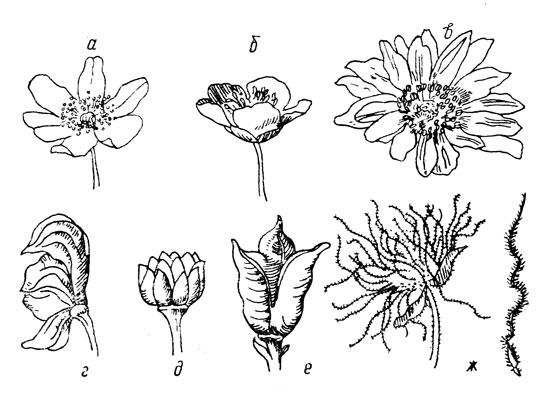
Familie der Hahnenfußgewächse (Ranunculaceae). Die Blütenstruktur dieser Pflanzen ist vielfältiger als die der vorherigen Familie (Abb. 9). In einigen Gattungen haben die Blüten eine radiale Symmetrie (aktinomorph) – Butterblume, Pfingstrose, Anemone, während sie in anderen eine einseitige Symmetrie (zygomorph) haben, wie zum Beispiel beim Rittersporn. Einige haben sowohl einen Kelch als auch eine Krone, andere haben eine einfache Blütenhülle, andere haben einen gekrönten Kelch und die Blütenblätter der Krone werden in Nektarien umgewandelt. Es gibt viele Staubblätter. Der Stempel besteht meist aus vielen Fruchtblättern, die jeweils einzeln geschlossen sind, und daher ist die Frucht dieser Pflanzen zusammengesetzt. Oberer Eierstock. Die Frucht ist modular aufgebaut und die einzelnen kleinen Früchte sind trocken, geschlossen oder blasig (selten ist die Frucht saftig – Actaea).
Familie der Rosaceae (Rosaceae). Die Blüte ist polysymmetrisch – korrekt, nach Typ fünf angeordnet – (Abb. 10); der Kelch besteht aus fünf verwachsenen Kelchblättern (manche haben auch einen Unterkelch aus fünf Kelchblättern); unverwachsene Krone aus fünf Blütenblättern (selten aus 4, 6, 8 Blütenblättern); viele Staubblätter (selten 5 oder 4); Stempel, bestehend aus einem, fünf oder mehreren Fruchtblättern; die Position des Eierstocks gilt als niedriger; Die Frucht ist eine Steinfrucht (Kirsche, Pflaume, Pfirsich), vorgefertigt (Himbeere, Brombeere), falsch (Erdbeere, Wildrose) oder halbfalsch (Birne, Apfel).

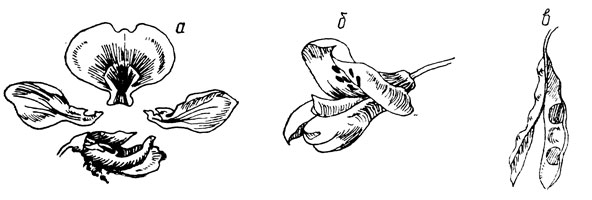
Mottenfamilie (Papilionaceae). Betrachtet man die Blüte im Profil, ähnelt sie einem sitzenden Schmetterling. Die Blüte ist nach Typ fünf einseitig symmetrisch angeordnet (Abb. 11). Die fünf Kelchblätter bilden einen sitzenden Kelch in Form einer mehr oder weniger langen Röhre. Die Krone besteht aus einem, meist größeren Blütenblatt – einem Segel (Flagge), zwei gleich großen seitlichen Blütenblättern – Flügeln und zwei in ihrem oberen Teil verwachsenen Blütenblättern – Booten. Es gibt 10 Staubblätter, von denen neun Filamente zusammenwachsen und das zehnte frei bleibt, oder die Filamente aller zehn Staubblätter wachsen zusammen und bilden eine Röhre. Der Stempel besteht aus einem einzelnen Fruchtblatt mit einem oberen Fruchtknoten. Die Frucht ist eine Bohne (Hülsenfrucht), die sich entlang zweier Nähte öffnet oder sich in einsamige oder nicht öffnende Lappen (Erdnuss) aufteilt.
Familie der Doldenblütler. Fast alle Vertreter dieser Familie haben den gleichen Blütenaufbau (Abb. 12), meist strahlend symmetrisch (aktinomorph). Der Kelch besteht aus fünf Zehen, die jedoch bei den meisten Arten fehlen. Krone aus fünf Blütenblättern, nicht verwachsen, unterschiedlich gefärbt. Bei einigen Arten sind die äußeren Blütenblätter größer; Staubblätter 5, sie befinden sich zwischen den Blütenblättern der Krone. Der Stempel besteht aus zwei Fruchtblättern mit zwei Säulen und einem unteren Fruchtknoten. Die Frucht ist trocken und spaltet sich meist in zwei einsamige Ventile auf (bei Koriander usw. bleibt sie intakt).

Familie der Lippenblütengewächse (Labiatae). Die Struktur der Blüte bei Vertretern dieser Familie ist spezieller und typischer (Abb. 13). Die Blüte ist monosymmetrisch. Der Kelch besteht aus fünf zu einer regelmäßigen Röhre verwachsenen Kelchblättern mit fünf Zähnen; nur bei einigen ist es zweilippig. Die Blütenkrone besteht aus fünf Blütenblättern, die zu einer Röhre verschmolzen sind, im oberen Teil jedoch frei sind und eine längere oder kürzere Oberlippe (erhalten aus der Verschmelzung zweier Blütenblätter) und eine Unterlippe (erhalten aus der Verschmelzung der verbleibenden drei Blütenblätter) bilden. . Staubblätter 4 oder 2 (selten); wenn sie zu viert sind, sind zwei davon länger und zwei kürzer (doppelte Stärke – Didynamus). Der Stempel besteht aus zwei Fruchtblättern, der Fruchtknoten ist oben; Später entsteht jedoch eine zusätzliche Trennwand, wodurch die Frucht im reifen Zustand trocken wird und in vier einkernige Nüsse zerfällt. Diese Familie zeichnet sich durch zwei weitere typische vegetative Merkmale aus – einen tetraedrischen Stamm und entgegengesetzt angeordnete Blätter. Fast alle Arten dieser Familie sind reich an ätherischen Ölen.

Familie norichnikovye (Scrophulariaceae). Einige Arten dieser Familie ähneln hinsichtlich der Struktur der Blüte den Arten der vorherigen Familie. Allerdings gibt es hier eine größere Vielfalt in der Struktur der gesamten Blüte (mit Ausnahme des Stempels). Es überwiegen monosymmetrische (zygomorphe) Blüten (Abb. 14). Der Kelch besteht aus vier bis fünf verwachsenen Kelchblättern. Blütenkrone – ebenfalls aus vier oder fünf Blütenblättern, verwachsen, ganz unterschiedlich angeordnet (polysymmetrisch und monosymmetrisch, mit oder ohne Sporn, offen oder zweilippig geschlossen usw.). Staubblätter 5, 4 oder 2. Der Stempel besteht aus zwei Fruchtblättern, der Fruchtknoten ist oben, die Frucht ist eine Schachtel, die meist an zwei Nähten geöffnet ist, oder eine Schachtel, aus der die Samen durch Löcher heraustreten.

Familie der Korbblütler (Compositae). Obwohl die Blüten zahlreicher Arten (mehr als 20.000) dieser Familie eine recht einheitliche Struktur aufweisen – einige sind röhrenförmig und andere schilfförmig – zeichnen sie sich typischerweise durch eine besondere Struktur des Blütenstandes aus, die als Korb bezeichnet wird (Abb. 15). ). Am Boden des Korbes befinden sich die Blumen. Außen ist der Korb mit einer mehr oder weniger großen Anzahl grüner Blätter bedeckt (bedeckt), die seine Hülle (Involucrum) bilden. Die Blüten dieses Blütenstandes verschiedene Typen und die Gattungen dieser Familie sind entweder nur röhrenförmig oder nur rohrförmig, oder die Randblüten sind rohrförmig und die mittleren röhrenförmig. Im Allgemeinen sind beide jedoch wie folgt angeordnet: Es gibt keinen Kelch, und stattdessen entwickeln einige Haare (die die sogenannte Fledermaus oder das Büschel des Fötus bilden), Schuppen oder andere Fortsätze; Die Blütenkrone ist immer entweder vollständig röhrenförmig und besteht aus fünf verwachsenen Blütenblättern, oder sie ist an der Basis röhrenförmig und wird an der Spitze liguliert; fünf Staubblätter, die mit ihren Staubbeuteln (Synantherea) zu einer Röhre verwachsen sind, durch die die Stempelsäule verläuft; der Stempel besteht aus zwei Fruchtblättern, der Fruchtknoten ist unterständig; Die Frucht ist ein Fruchtsamen (Achena oder Achenium). Ein Nichtfachmann kann einen Blütenstandkorb als eine gewöhnliche Blume betrachten, indem er die Hülle des Korbes für einen Kelch, die am Rand des Korbes befindlichen Schilfrohrblüten für die Blütenblätter der Krone und die röhrenförmigen Blüten für die Staubgefäße hält. Alle Pflanzen dieser Familie, deren Blüten nur Schilf sind, bilden die Unterfamilie Liguliflorae, und diejenigen Arten, deren Blüten nur röhrenförmig oder röhrenförmig (Median) und Schilf (Rand) sind, bilden die Unterfamilie Tubuliflorae.
Das Ablegen der Blüten erfolgt bei Sojabohnen in den frühen Wachstumsphasen. Je kürzer die Vegetationsperiode einer Sorte ist, desto früher bilden sie sich. Sehr frühe Sorte Nördlicher Rekord beginnt in der Phase zweier echter Blätter mit dem Blumenlegen. Sorte für die Zwischensaison Primorskaja 529 Sie bilden sich viel später – bei Vorhandensein von 5–7 Blättern, und die späte japanische Form – erst bei der Entwicklung von 9–10 Blättern. Nach dem Legen der embryonalen Blütenknollen erfolgt nach 3–6 Tagen deren Differenzierung in sexuelle Elemente. Vielfalt Nördlicher Rekord dies wird in der Phase der drei echten Blätter festgestellt.
Wenn wir das Erscheinen der ersten Blütenknollen als das Ende des Lichtstadiums betrachten, beträgt seine Dauer in verschiedenen Formen (je nach Vegetationsperiode, Sorte und meteorologischen Bedingungen) am natürlichen Tag 18–40 Tage.
Soja gehört zur Gruppe der Selbstbestäuber. Die natürliche Hybridisierung beträgt 0,11–0,16 %. Die Fremdbestäubung bei Sojabohnen beträgt 0,5 %.
Natürliche Hybriden unter dem Einfluss meteorologischer Faktoren können manchmal häufiger als üblich auftreten. Die Anpassungsfähigkeit der Sojabohne an die Selbstbestäubung wird durch die Struktur der Blüte und den gesamten Verlauf des Bestäubungsprozesses bestätigt. Vom Zeitpunkt des Erscheinens der Knospe bis zur Entwicklung des Fruchtknotens, in dem die Samen bereits sichtbar sind, lassen sich acht Phasen feststellen (Abb. 1).
Reis. 1. Schema der Dynamik der Blütenentwicklung (Bedeutung der Zahlen siehe Text)
Während Erste Und zweite Phasen Wenn die Blütenkrone geschlossen ist und sich unterhalb der Kelchzähne befindet, ist die Vorbereitung der Narbe und der Staubblätter für die Bestäubung abgeschlossen. Die Reifung und Bestäubung der Staubblätter erfolgt, wenn die Blütenkrone noch fest verschlossen ist und sich etwa auf Höhe der Kelchzähne oder etwas höher befindet ( Phase 3 Und 4). Pollen Aus den Staubbeuteln ergießt es sich auf die Narbe, bedeckt mit einer klebrigen Masse. Die Öffnung der Blütenkrone erfolgt etwa 15–20 Minuten nach der Pollenkeimung. Daher öffnet sich die Blütenkrone nach Abschluss der Bestäubung und bis sie vollständig geöffnet ist ( Phase 6) beginnt sich die Narbe merklich zu verformen. Während der Welkezeit der Blütenkrone wird ein verstärktes Wachstum des Eierstocks beobachtet. Bei mangelnder Wärme öffnen sich die Blüten möglicherweise überhaupt nicht, was ihre Bestäubung nicht beeinträchtigt.
Sojabohnenblüten sehr klein, geruchlos, in der Blattmasse kaum wahrnehmbar. Zunächst erscheinen einzelne Blüten, die sich am Hauptstamm im unteren oder mittleren Teil befinden, in den Achseln des 2.–10. Blattes. Nach 4–6 Tagen beginnt in der gesamten Pflanze eine kräftige Blüte, die 5 bis 11 Tage anhält. Dann verlangsamt sich der Blüteprozess. Die Blütephase reicht je nach Frühreife der Sorte aus Bevorzugte Umstände dauert 12 bis 43 Tage. Vom Erscheinen der Knospe bis zum vollständigen Öffnen der Blüten vergehen etwa 3 bis 4 Tage und vom Öffnen der Blüte bis zum vollständigen Welken der Blütenkrone etwa die gleiche Zeit. Eine angenehme Blüte und Bohnenbildung wird durch eine Lufttemperatur von 20-26 °C erleichtert. Sehr trockenes, heißes oder kaltes Wetter verlangsamt die Blütenentwicklung. Kalte Nächte, auch bei ausreichend hohen Tagestemperaturen, beeinträchtigen den Blüteprozess. Der negative Einfluss der Nachttemperatur auf die Blütenentwicklung ist darauf zurückzuführen, dass die Bildung der Geschlechtselemente hauptsächlich zu dieser Zeit stattfindet. Die Blüte gelingt gut bei einer relativen Luftfeuchtigkeit von 65-80 %. Eine sehr hohe Luftfeuchtigkeit führt zu einer Hemmung der Blüte und verursacht das, was wir in manchen Jahren beobachtet haben. Nach chinesischen Angaben wird die Blüte durch eine Luftfeuchtigkeit von 74-80 % begünstigt, ab 90 % kommt es nicht mehr dazu. Durch die Anwendung können Sie den Verlauf der Blüte beeinflussen Chemikalien und Störung der normalen Abfolge der Blattentwicklung. So zum Beispiel die Blüte von Sojabohnen in der Zwischensaison Lincoln und spät Ogden An einem langen Tag wurde es durch das Besprühen der Pflanzen mit Auxin beschleunigt und es erschien eine beträchtliche Anzahl von Blüten in den unteren Internodien.
Sojabohnen blühen nachts nicht. Am energischsten ist dieser Vorgang morgens. Je nach Gebiet und meteorologischen Bedingungen ist die Blüte zu unterschiedlichen Zeiten zur Bestäubung bereit. Das Massenknacken der Staubbeutel dauert 2-3 Stunden. Je mehr Pollen auf die Narbe gelangen, desto kräftiger keimt sie.