Почему дети похожи на своих родителей? Почему в некоторых семьях распространены определенные заболевания, такие как дальтонизм, полидактилия, гипермобильность суставов, муковисцидоз? Почему есть ряд болезней, которыми болеют только женщины, а другими - только мужчины? Сегодня все мы знаем, что ответы на эти вопросы нужно искать в наследственности, то есть хромосомах, которые ребенок получает от каждого из родителей. И этим знанием современная наука обязана Томасу Ханту Моргану - американскому генетику. Он описал процесс передачи генетической информации и разработал вместе с коллегами хромосомную теорию наследования (ее часто называют хромосомная теория Моргана), которая стала краеугольным камнем современной генетики.
История открытия
Неправильно было бы говорить, что Томас Морган был первым, кто заинтересовался вопросом передачи генетической информации. Первыми исследователями, которые пытались понять роль хромосом в наследовании, можно считать работы Чистякова, Бенедена, Рабля в 70-80-х годах XIX века.
Тогда не было микроскопов настолько мощных, чтобы можно было разглядеть хромосомные структуры. Да и самого термина "хромосома" тогда тоже не было. Его ввел немецкий ученый Генрих Вальдейр в 1888 году.
Немецкий биолог Теодор Бовери в результате своих экспериментов доказал, что для нормального развития организма ему необходимо нормальное для его вида количество хромосом, а их избыток или нехватка ведет к тяжелым порокам развития. Со временем его теория блестяще подтвердилась. Можно сказать, что хромосомная теория Т. Моргана получила свою отправную точку именно благодаря исследованиям Бовери.
Начало исследований
Обобщить существующие знания о теории наследственности, дополнить и развить их сумел Томас Морган. В качестве объекта для своих экспериментов он выбрал плодовую мушку, и не случайно. Это был идеальный объект для исследований передачи генетической информации - всего четыре хромосомы, плодовитость, небольшая продолжительность жизни. Морган начал проводить исследования, используя чистые линии мушек. Вскоре он обнаружил, что в половых клетках одинарный набор хромосом, то есть 2 вместо 4. Именно Морган обозначил женскую половую хромосому как Х, а мужскую - как Y.
Сцепленное с полом наследование
Хромосомная теория Моргана показала, что существуют определенные признаки, сцепленные с полом. Мушка, с которой ученый проводил свои опыты, в норме имеет красный цвет глаз, но в результате мутации этого гена в популяции появились белоглазые особи, причем среди них было гораздо больше самцов. Ген, который отвечает за цвет глаз мушки, локализуется на Х-хромосоме, на У-хромосоме его нет. То есть при скрещивании самки, на одной Х-хромосоме которой есть мутировавший ген, и белоглазого самца вероятность наличия этого признака у потомства будет связана с полом. Проще всего показать это на схеме:
- P: ХХ" х Х"У;
- F 1: ХХ", ХУ, Х"Х", Х"У.
Х - половая хромосома самки или самца без гена белых глаз; Х"- хромосома с геном белых глаз.
Расшифруем результаты скрещивания:
- ХХ" - красноглазая самка, носитель гена былых глаз. За счет наличия второй Х-хромосомы этот мутировавший ген "перекрывается" здоровым, и в фенотипе признак не проявляется.
- Х"У - белоглазый самец, который получил от матери Х-хромосому с мутировавшим геном. За счет наличия только одной Х-хромосомы мутантный признак нечем перекрыть, и он появляется в фенотипе.
- Х"Х" - белоглазая самка, унаследовавшая по хромосоме с мутантным геном от матери и отца. У самки только если обе Х-хромосомы несут в себе ген белых глаз, он проявится в фенотипе.
Томаса Моргана пояснила механизм наследования многих генетических заболеваний. Поскольку на Х-хромосоме гораздо больше генов, чем на У-хромосоме, понятно, что она отвечает за большинство признаков организма. Х-хромосома от матери передается как сыновьям, так и дочерям, вместе с генами, отвечающими за свойства организма, внешние признаки, заболевания. Наряду с Х-сцепленным существует У-сцепленное наследование. Но У-хромосома есть только у мужчин, потому если в ней происходит какая-либо мутация, она может передаться только потомком мужского пола.
Хромосомная теория наследственности Моргана помогла понять закономерности передачи генетических заболеваний, однако трудности, связанные с их лечением, не разрешены до сих пор.
![]()
Кроссинговер
В ходе исследований учеником Томаса Моргана Альфредом Стертевантом было открыто явление кроссинговера. Как показали дальнейшие опыты, благодаря кроссинговеру появляются новые комбинации генов. Именно он нарушает процесс сцепленного наследования.
Таким образом хромосомная теория Т. Моргана получила еще одно важное положение - между происходит кроссинговер, а его частота определяется расстоянием между генами.
Основные положения
Чтобы систематизировать результаты опытов ученого, приведем основные положения хромосомной теории Моргана:
- Признаки организма зависят от генов, заложенных в хромосомах.
- Гены одной хромосомы передаются потомству сцепленно. Сила такого сцепления тем больше, чем меньше расстояние между генами.
- В гомологичных хромосомах наблюдается явление кроссинговера.
- Зная частоту кроссинговера определенной хромосомы, можно вычислить расстояние между генами.

Второе положение хромосомной теории Моргана также называется правилом Моргана.
Признание
Результаты исследований были восприняты блестяще. Хромосомная теория Моргана стала прорывом в биологии ХХ века. В 1933 году за открытие роли хромосом в наследственности ученому была присуждена Нобелевская премия.
Еще через несколько лет Томас Морган получил медаль Копли за выдающиеся достижения в области генетики.
Сейчас хромосомная теория наследственности Моргана изучается в школах. Ей посвящено множество статей и книг.
Примеры сцепленного с полом наследования
Хромосомная теория Моргана показала, что свойства организма определяются заложенными в нем генами. Фундаментальные результаты, которые получил Томас Морган, дали ответ на вопрос о передаче таких заболеваний, как гемофилия, синдром Лоу, дальтонизм, болезнь Бруттона.

Оказалось, что гены всех этих болезней располагаются на Х-хромосоме, и у женщин эти заболевания проявляются гораздо реже, так как здоровая хромосома может перекрыть хромосому с геном болезни. Женщины, не зная об этом, могут быть носителями генетических болезней, которые потом проявляются у детей.
У мужчин Х-сцепленные заболевания, или фенотипические признаки, проявляются, поскольку нет здоровой Х-хромосомы.
Хромосомная теория наследственности Т. Моргана применяется при анализе семейных анамнезов на предмет генетических заболеваний.
Работы Моргана заложили основы хромосомной теории наследственности, они показали, что ограничения в свободной комбинаторике некоторых генов обусловлены расположением этих генов в одной хромосоме и их физическим сцеплением.
Морганом было установлено, что сцепление генов, расположенных в одной хромосоме, не является абсолютным. Во время мейоза хромосомы одной пары могут обмениваться гомологичными участками между собой с помощью процесса, который называется кроссинговером. Чем дальше друг от друга расположены гены в хромосоме, тем чаще они разделяются кроссинговером. На основе этого феномена была предложена мера силы сцепления генов - процент кроссинговера - и построены первые генетические карты хромосом для разных видов дрозофилы.
В качестве объекта генетического анализа была выбрана плодовая мушка дрозофила и Морган изучал наследование у нее разных признаков.
Скрестив гомозиготную самку с серыми телом и длинными крыльями (домин), с гомозиготным чернокрылым короткокрылым самцом, в F1 – однообразие (серое тело, длинные крылья)
Оказалось, что результаты будут разные в зависимости от пола гибрида.
Если гибридным был самец, то в потомстве получалось 2 фенотипических класса полностью повторяющих признаки родителей.
Если гибридной была самка, то получалось 4 фенотипических классов потомком в неравных пропорциях. Большую часть потомства (83%) составляют потомки с родительскими признаками, меньшую (17%) – особи с новыми комбинациями признаков.
Морган сделал вывод, что сцепление может быть неполным, где группа сцепления нарушается кроссинговером.
Необычность процентного соотношения у потомков объясняется тем, что кроссинговер происходит не всегда, частота кроссинговера зависит от расстояния между генами – чем больше расстояние, тем меньше силы сцепления между генами, тем чаще кроссинговер.
Гаметы, в которые попали хромосомы, не прошедшие кроссинговер, называются некроссоверные.
Если в гаметах хромосомы претерпевшие кроссинговер – кроссоверные.
6. Основные положения хромосомной теории наследственности
1. Гены расположены в хромосомах линейно в определенных участках – локусах. Аллельные гены занимают одинаковые локусы гомологичных хромосом.
2. Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе или сцеплено. Число групп сцепления = числу хромосом в гаплоидном наборе.
3. Между гомологичными хромосомами возможен кроссинговер, нарушающий сцепление
4. процесс кроссинговера прямо пропорционален расстоянию между генами.
1% кроссинговера = 1 сантиморганида
7. Понятие о цитоплазматической наследственности
Наличие некоторого количества наследственного материала в цитоплазме в виде кольцевых молекул ДНК митохондрий и пластид, а также других внеядерных генетических элементов дает основание специально остановиться на их участии в формировании фенотипа в процессе индивидуального развития.
Цитоплазматические гены не подчиняются менделевским закономерностям наследования, которые определяются поведением хромосом при митозе, мейозе и оплодотворении. В связи с тем что организм, образуемый вследствие оплодотворения, получает цитоплазматические структуры главным образом с яйцеклеткой, цитоплазматическое наследование признаков осуществляется по материнской линии. Такой тип наследования был впервые описан в 1908 г. К. Корренсом в отношении признака пестрых листьев у некоторых растений.
Формирование хромосомной теории.В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бо-вери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911-1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом (рис. 3.10). В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная величина кроссинговера не превышает 50%. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
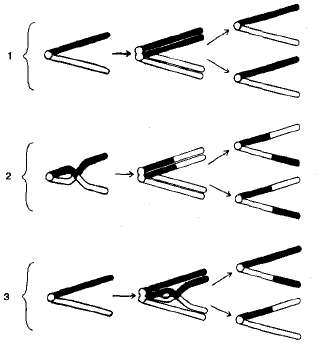
Рис. 3.10. Схема кроссинговера: I - отсутствие кроссинговера; 2 - кроссинговер на стадии двух хромосом; 3 - кроссинговер на стадии четырех хроматид.
Рассмотрим один из первых экспериментов Т. Моргана по изучению сцепленного наследования. При скрещивании дрозофил, различающихся по двум парам сцепленных признаков -серых с зачаточными крыльями и черных с нормальными крыльями - гибриды F 1 были серыми с нормальными крыльями (рис. 3.11).
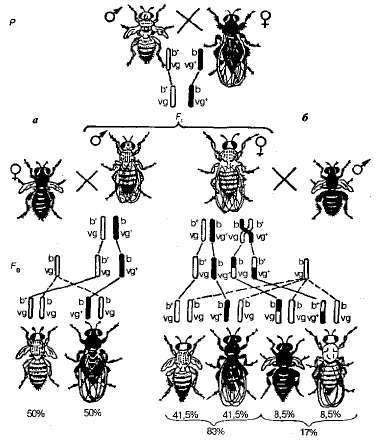
Рис. 3.11 . Наследование сцепленных признаков у дрозофилы: а-полное сцепление (дигетерозигртен самец, у которого отсутствует кроссинговер); б - сцепление с кроссинговером {дигетерозигатт самка, у которой кроссинговер не подавлен); b + ,b - ~ серая и черная окраски тела; vg + , vg - - нормальные и зачаточные крылья соответственно.
Далее были проведены два типа анализирующих скрещиваний. В первом из них брали дигетерозиготных самцов F 1 , и скрещивали с гомозиготными по рецессивным аллелям самками, а во втором - дигетерозиготную самку скрещивали с самцами, рецессивными по обоим признакам (черное тело и зачаточные крылья). Результаты этих скрещиваний оказались разными (см. рис. 3.11).
В первом случае были получены потомки с исходными для этого опыта родительскими (Р) фенотипами, т. е. серые мухи с зачаточными крыльями и черные мухи с нормальными крыльями в отношении 1:1. Следовательно, данная дигетерозигота образует только два типа гамет (b + vg и bvg +) вместо четырех. Исходя из указанного расщепления, можно предположить, что у самца наблюдается полное сцепление генов.
Во втором случае в F 2 наблюдалось иное расщепление. Кроме родительских комбинаций признаков, появились новые - мухи с черным телом и зачаточными крыльями, а также с серым телом и нормальными крыльями. Правда, количество рекомбинантных потомков невелико и составляет 17%, ародительских - 83%. Причиной появления небольшого количества мух с новыми сочетаниями признаков является кроссинговер, который приводит к новому рекомбинантному сочетанию аллелей генов b и vg в гомологичных хромосомах. Эти обмены происходят с вероятностью 17% и в итоге дают два класса рекомбинантов с равной вероятностью - по 8,5%.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость , которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Понятие о генетической карте. Т. Морган и его сотрудники К. Бриджес, А. Стертеванти Г. Меллер экспериментально показали, что знание явлений сцепления и кроссинговера позволяет не только установить группу сцепления генов, но и построить генетические карты хромосом, на которых указаны порядок расположения генов в хромосоме и относительные расстояния между ними.
Генетической картой хромосом называют схему взаимного расположения генов, находящихся в одной группе сцепления. Такие карты составляются для каждой пары гомологичных хромосом.
Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи , аспергилл), для бактерий и вирусов.
Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что теперь широко используется в селекционной практике. Так, создание штаммов микроорганизмов, способных синтезировать необходимые для фармакологии и сельского хозяйства белки , гормоны и другие сложные органические вещества, возможно только на основе методов генной инженерии, которые, в свою очередь, базируются на знании генетических карт соответствующих микроорганизмов.
Генетические карты человека также могут оказаться полезными в здравоохранении и медицине. Знания о локализации гена в определенной хромосоме используются при диагностике ряда тяжелых наследственных заболеваний человека. Уже теперь появилась возможность для генной терапии, т. е. для исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов способствует также пониманию эволюционного процесса.
Основные положения хромосомной теории наследственности. Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
- Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
- Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
- Гены расположены в хромосоме в линейной последовательности.
- Гены одной хромосомы образуют группу сцепления, благодаря нему происходит сцепленное наследование некоторых признаков. При этом сила сцепления находится в обратной зависимости от расстояния между генами.
- Каждый биологический вид характеризуется определенным набором хромосом - кариотипом.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"
Создателем хромосомной теории (ХТ) является учёный Томас Морган. ХТ является результатом изучения наследственности на клеточном уровне.
Суть хромосомной теории :
Материальными носителями наследственности являются хромосомы.
Основными доказательством этому является:
Цитогенетический параллелизм
Хромосомное определение пола
Сцепленное с полом наследование
Сцепление генов и кроссинговер
Основные положения хросомной теории:
Наследственные задатки (гены) локализованы в хромосомах.
Гены расположены в хромосоме в линейном порядке.
Каждый ген занимает определенный участок (локус). Аллельные гены занимают аналогичные локусы в гомологичных хромосомах.
Гены, локализованные в одной хромосоме, наследуются совместно, сцеплено (Закон Моргана) и образуют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом (n).
Между гомологичными хромосомами возможен обмен участками, или рекомбинация.
Расстояние между генами измеряются в процентах кроссинговера – морганидах.
Частота кроссинговера обратно пропорциональна расстоянию между генами, а сила сцепления между генами обратно пропорциональна расстоянию между ними.
Цитогенетический параллелизм
Дипломником Моргана Сюттоном было замечено что поведение генов по Менделю, совпадает с поведение хромосом: (ТАБЛИЦА – цитогенетический паралелизм)
Каждый организм несёт 2-а наследственных задатка, в гамету входит только 1- ин наследственный задаток из пары. При оплодотворении в зиготе и далее в организме опять 2-а наследственных задатка по каждому признаку.
Точно так же ведут себя и хромосомы, что можно предположить что гены лежат в хромосомах и наследуются вместе с ними.
Хромосомное определение пола
В 1917 году Алленом было показано что мужские и женские особи у мхов отличаются по набору хромосом. В клетках диплоидной ткани мужского организма половые хромосомы XиY, в женскомXиX. Таким образом Хромосомы определяют такой признак как пол, а следовательно могут быть материальными носителями наследственности. Позже хромосомное определение пола было показано и для других организмов, в том числе и для человека.(ТАБЛИЦА)
Сцепленное с полом наследование
Поскольку половые хромосомы различны у мужских и женских организмов, признаки, гены которых, расположены в Х или Yхромосомах, будут наследовать по-разному. Такие признаки называютсясцепленными с полом признаками .
Особенности наследования сцепленных с полом признаков
Не соблюдается 1 закон Менделя
Реципрокные скрещивания дают разный результат
Имеет место крисс-кросс (или наследование крест-накрест).
Впервые наследование связанное с признаком было обнаружено Морганом у дрозофилы.
|
W + -красные глаза |
(C) X W+ X W+ * X w Y |
(C) X w X w * X W+ Y |
|||||
|
w – белые глаза | |||||||
|
(CЖ)X W + X w – Красные глаза |
X w X W + - Красные глаза |
||||||
|
(CМ)X W + Y– Красные глаза |
X w Y– Белые глаза |
||||||
|
Таким образом наследование выявленной Морганом мутация – “белые глаза” - white, характеризовалась перечисленными выше особенностями: Закон единообразия несоблюдался В 2-ух реципрокных скрещиваниях получено разное потомство Во втором скрещивании сыновья получают признак матери (белые глаза), дочери – признак отца (красные глаза). Такое наследование и называется «наследование крисс-кросс» |
|||||||
(ТАБЛИЦА сцепленное с полом наследование )
Сцепленное с полом наследование объясняется отсутствием в Yхромосоме генов, аллельных генамXхромосоме.Yхромосома намного меньше Х хромосомы, в ней, в настоящее время, локализовано 78(?) генов, в то время как вXхромосоме их более 1098.
Примеры сцепленных с полом наследований:
Гемофилия, дистрофия Дюшенна, синдром Данкана, синдром Альпорта, и др.
Есть гены, которые наоборот содержатся в Yхромосоме и отсутствуют вXхромосоме, они, следовательно, встречаются только в мужских организмах, и никогда в женских (голандрическое наследование) и передаются только сыновьям от отца.
Сцепление генов и кроссинговер
В генетике было известно такое явления как «притяжение генов»: некоторые неаллельные признаки наследовались не независимо, как должны по IIIзакону Менделя, а наследовались вместе, не давали новых комбинаций. Морган объяснил это тем, что эти гены находятся в одной хромосоме, поэтому они расходятся в дочерние клетки вместе одной группой, как бы сцеплено. Он назвал это явление –сцепленным наследованием .
Закон сцепления Моргана:
Гены расположенные в одной хромосоме наследуются совместно, сцеплено.
Гены расположенные в одной хромосоме образуют группу сцепления. Число групп сцепления равно «n» - гаплоидному числу хромосом.
Скрещивали гомозиготные линии мух с серым цветом тела и длинными крыльями и мух, имеющих чёрное тело и короткие крылья. Гены цвета тела и длинны крыльев – сцеплены, т.е. лежат в одной хромосоме.
|
А- серое тело а- чёрное тело B- нормальные крылья (длинные) b- зачаточные крылья |
(С Ж) AABBxaabb(CМ) |
|||||
|
Серые длиннокрылые |
Чёрные короткокрылые |
|||||
|
Запись в хромосомном выражении |
||||||
|
Серое тело Длинные крылья |
Чёрное тело Короткое тело |
|||||
|
|
|
|||||
|
Все мухи имеют серое тело и длинные крылья |
||||||
|
Т.е. в
этом случае закон единообразия
гибридов Iпоколения
соблюдается. Однако вF 2 вместо ожидаемого расщепления 9:3:3:1
получилось отношение на 3 серых
длиннокрылых на 1 часть чёрных
короткокрылых, т.е. новых сочетаний
признаков не появлялось. Морган
предположил что дегетерозиготыF 2 - ( |
||||||
|
|
||||||
|
Серое тело Длинные крылья |
Чёрное тело Короткое тело |
|||||
|
|
|
|||||
|
F a |
|
|
||||
|
Серое тело Длинные крылья |
Чёрное тело Короткие крылья |
|||||
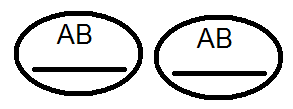

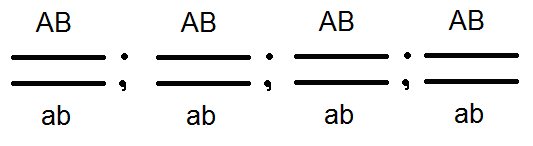
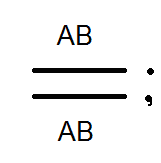
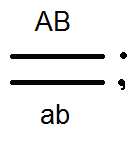 )продуцируют
(дают) гаметы не 4, а только 2 типов -
)продуцируют
(дают) гаметы не 4, а только 2 типов - и
и .
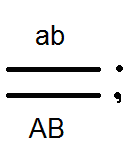
Проведенные анализирующие скрещивания
это подтвердило:
.
Проведенные анализирующие скрещивания
это подтвердило: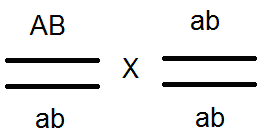
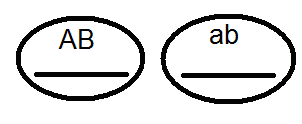
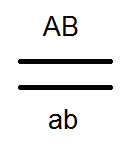

В результате в F 2 расщепление идёт как при моногибридном скрещивании 3:1.
|
|
||||
|
|
|
|||
|
|
|
|
|
|
|
Серое тело Длинные крылья |
Серое тело Длинные крылья |
Серое тело Длинные крылья |
Чёрное тело Короткие крылья |
|



Кроссинговер.
В небольшом проценте случаев в F 2 в опытах Моргана появлялись мухи с новыми сочетаниями признаков: крылья длинные, тело черное; крылья короткие, а тело серое. Т.е. признаки «расцепились». Морган объяснил это тем, что хромосомы во время конъюгации в мейозе обмениваются генами. В результате получаются особи с новыми сочетаниями признаков, т.е. как и положено по третьему закону Менделя. Морган назвал этот обмен генами рекомбинацией.
Позже цитологи действительно подтвердили гипотезу Моргана, обнаружив обмен участками хромосом у кукурузы и у саламандры. Они назвали этот процесс кроссинговер.
Кроссинговер увеличивает разнообразие потомства в популяции.
После того, как в генетике утвердилось понятие о наследственных факторах, были проведены исследования с целью определить, с какими клеточными структурами они связаны.
Факты, установленные генетическими и цитологическими работами еще в начале текущего столетия, показывали, что носители наследственных факторов (генов) - хромосомы.
В результате дальнейшего развития генетики появилась хромосомная теория наследственности . Создатель ее - американский генетик Т. Морган.
Ученый проводил исследования на плодовой мушке дрозофиле, которую можно легко разводить в пробирках. У этой мушки очень короткий цикл развития: в течение двух недель из оплодотворенного яйца через промежуточные стадии личинки и куколки развивается взрослая особь, способная сразу же давать потомство. Одна оплодотворенная самка может дать несколько сот новых насекомых.
Дрозофила имеет большое число хорошо отличимых признаков, наследование которых легко наблюдать при различных видах скрещиваний. В соматических клетках у нее всего четыре пары хромосом.
Вследствие названных особенностей дрозофила оказалась очень удобным объектом для генетических исследований. На основе опытов с ней были разработаны многие важнейшие вопросы общей генетики.
Многочисленные лабораторные опыты, проведенные Т. Морганом, показали, что гены, находящиеся в одной хромосоме, наследуются, как правило, совместно, то есть оказываются сцепленными и поэтому не подчиняются установленному Г. Менделем правилу независимого комбинирования.
В одном из опытов Т. Моргана дрозофила, имевшая серую окраску тела и длинные крылья, была скрещена с особью, имевшей черную окраску тела и рудиментарные (укороченные) крылья. Первое поколение мух было с серым телом и длинными крыльями. При скрещивании этих гибридов между собой в F 2 не произошло независимого распределения признаков по двум аллельным парам (серое тело - черное тело, длинные крылья - рудиментарные крылья) в отношении 9:3:3:1. Среди гибридов F 1 преобладающее число особей унаследовало такую же комбинацию признаков, какой она была у родительских форм (серые длиннокрылые и черные короткокрылые), и лишь очень небольшая часть мух была с перекомбинированными признаками (серые короткокрылые и черные длиннокрылые). Этот пример показывает, что гены, обусловливающие признаки серого тела и длинных крыльев и черного тела и коротких крыльев, наследуются преимущественно вместе.
На основании этого и большого числа подобных опытов Т. Морган пришел к выводу, что материальная основа сцепления генов - хромосома. Каждая из хромосом по своей длине неоднородна, она состоит из отдельных элементарных наследственных единиц - генов. У любого вида организмов их всегда во много раз больше, чем хромосом. Следовательно, в каждой хромосоме находится определенное число генов, которые наследуются совместно, образуя, так называемые группы сцепления. Число групп сцепления соответствует числу пар гомологичных хромосом.
Изучая явление сцепления генов, Т. Морган и его ученики установили, что сцепление почти никогда не бывает полным.
В разбираемом примере оно также не было полным, поскольку в небольшом числе случаев отмечена перекомбинация генов. Если гены разных аллельных пар лежат в одной и той же хромосоме, то есть сцеплены, то единственной причиной их перекомбинации может быть процесс конъюгации гомологичных хромосом в профазе мейоза. Во время конъюгации парные хромосомы сближаются и прикладываются одна к другой гомологичными участками, образуя биваленты (четверки хроматид).
В это время между хроматидами может происходить обмен гомологичными участками. Этот процесс получил название перекреста хромосом или (от англ. кроссинг - перекрещивание).
Показана схема перекреста хромосом и рекомбинации находящихся в них генов. Две парные хромосомы в результате перекреста и последующего разрыва обмениваются участками. Два гена А к В, расположенные первоначально в одной хромосоме, в результате кроссинговера оказываются в разных хромосомах и попадают в разные гаметы.
Гаметы с хромосомами, претерпевшими кроссинговер, называются кроссоверными, а гаметы, образованные хромосомами без кроссинговера, - некроссоверными. Соответственно этому и особи, возникшие с участием кроссоверных гамет, называются кроссоверными, или рекомбинантными, а образованные без них - некроссоверными, или нерекомбинантными.
Рекомбинация генов в процессе скрещивания приводит к новообразованиям. Возникают гибридные формы, представляющие исходный материал для отбора и создания новых сортов растений и пород животных. Образование гибридных форм в природе дает материал для естественного отбора, поэтому имеет важнейшее значение в эволюции живых организмов.
Таким образом, перекомбинация генов в процессе мейоза осуществляется двумя путями - случайным расхождением негомологичных хромосом (правило независимого комбинирования по Г. Менделю) и процессом перекреста гомологичных хромосом (явление кроссинговера, установленное Т. Морганом).
В итоге разбора основных положений хромосомной теории наследственности можно сделать следующие выводы.
- Гены находятся в хромосомах, расположены линейно и образуют группу сцепления.
- Гены, локализованные в одной хромосоме, наследуются сцепленно; сила этого сцепления зависит от расстояния между генами.
- Между гомологичными хромосомами наблюдается перекрест, в результате которого происходит рекомбинация генов, имеющая важное значение как источник материала для естественного и искусственного отбора.
- Сцепление генов и их рекомбинация в результате перекреста - закономерные биологические явления, в которых выражается единство процессов наследственности и изменчивости организмов.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .