Хромосомное определение пола.
У раздельнополых организмов соотношение полов обычно составляют 1:1. Пол определяется половыми хромосомами. У раздельнополых организмов есть хромосомы, по которым самки и самцы не отличаются. Это аутосомы. Хромосомы, по которым самки и самцы отличаются друг от друга, называют половыми. Половую, палочковидную хромосому называют Х-хромосомой. Вторую половую, двуплечую хромосому называют У-хромосомой. Например, у человека 22 пары хромосом - аутосомы и пара половых хромосом. Пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы. У человека, дрозофилы и многих других животных женский пол гомогаметен (XX), т.е. яйцеклетки в отношении половых хромосом одинаковы. Мужской пол гетерогаметен (ХУ), т.е. развиваются сперматазоиды двух типов: либо с Х-, либо с У-хромосомой. При оплодотворении яйцеклетки спермием, несущим У- хромосому, будет развиваться мужской организм. У некоторых организмов другой тип определения пола, характеризующийся женской гетерогаметностью (например, у бабочек, птиц, пресмыкающихся). У домашних кур петух имеет набор хромосом XX, а курица ХУ.
На этой странице искали:
- хромосомное определение пола кратко
- хромосомное определение пола
У многих организмов соотношение между особями мужского и женского пола при изучении большого числа особей всегда примерно равное, т.е. расщепление по признаку пола проходит в отношении 1:1.
У плодовой мушки дрозофилы, на которой проведено множество генетических исследований, и у млекопитающих пол определяется следующим образом. В соматических клетках дрозофилы четыре пары хромосом. В число их входит три пары аутосом , т.е. хромосом, одинаковых у самца и самки, и одна пара хромосом, различных у особей мужского и женского пола. Это хромосомы, как было установлено, отвечают за наследование пола и поэтому были названы половыми хромосомами .
В клетках самок мух дрозофил имеются две одинаковые хромосомы, которые условно обозначают как Х-хромосомы. Следовательно, в диплоидных соматических клетках самки набор половых хромосом – ХХ. У самцов половые хромосомы отличаются от половых хромосом самок. В соматических клетках самца мухи дрозофилы имеется одна Х-хромосома и одна Y-хромосома. Поэтому набор половых хромосом самца обозначается XY. Следовательно, яйцеклетки женских организмов все одинаковы по хромосомному набору, т.к. в каждой из них имеются по одинаковому набору аутосом и одна половая хромосома. Все сперматозоиды также имеют по одному набору аутосом и одну половую хромосому, но половина сперматозоидов имеет Х-хромосому, а другая половина – Y-хромосому. Различаются они по набору генов, который в ней содержится. Поскольку гаметы с Х- и У-хромосом в результате мейоза образуются у самцов в равных количествах, то ожидаемое полов составляет 1:1, что совпадает с фактическими наблюдениями. Сходный способ определения полов присущим всем млекопитающим.
Наследование, сцепленное полом
В том случае, когда гены, ответственные за формирование признака, расположены в аутосомах, наследование осуществляется независимо от того, какой из родителей является носителем изучаемого признака. Однако ситуация резко изменяется, когда признаки определяются генами, лежащих в половых хромосомах. Черепаховая окраска, т.е. чередование черных и желтых пятен, встречается только у кошек. Котов с черепаховой окраской не бывает. Этот факт не могли объяснить, пока не стало известно, что наследование данного признака сцеплено с полом.
Рис. 32. Хромосомные комплексы самца и самки у дрозофилы
Рис. 33. Черепаховая окраска кошки

Черная окраска кошек определяется геном В, рыжая – геном b. Эти гены расположены в Х-хромосоме. В У-хромосоме они отсутствуют. Обозначим Х-хромосому, несущую аллель В, Х В, а Х-хромосому с аллелем b – Х b . Поэтому возможны такие комбинации: Х B X b – черная кошка; Х B У – черный кот X b – рыжая кошка; Х b У – рыжий кот Х B Х b – черепаховая кошка
Так как мужская особь имеет только одну Х-хромосому, то кот может быть или черным, или рыжим, но не может иметь черепаховую окраску, потому что для ее развития необходимо одновременное присутствие генов B и b.
У человека также известны признаки, сцепленные с полом. К ним относится, например, очень тяжелое наследственное заболевание гемофилия , при котором кровь теряет способность свертываться. У гемофиликов даже небольшие царапины и ссадины вызывают тяжелые кровотечения. Это заболевание встречается, за редчайшими исключениями, только у мужчин. Было установлено, что гемофилия обусловлена рецессивным геном, расположенным в Х-хромосоме, поэтому гетерозиготные по данному гену женщины обладают обычной свертываемостью крови. Рассмотрим, какое потомство может появиться у гетерозиготной женщины, вступившей в брак с нормальным по этому признаку мужчиной.
Ген, обуславливающий нормальную свертываемость крови, обозначим Н, а ген, при котором кровь теряет способность свертываться – h. Учитывая, что в генотипе женщины присутствуют две Х-хромосомы, а у мужчины – одна Х-хромосома и одна У-хромосома, запишем схему наследования гемофилии:
Женщина передает половине своих сыновей Х-хромосому с геном нормальной свертываемости крови, а половине – Х-хромосому с геном гемофилии. Среди ее сыновей могут быть и здоровые и гемофилики.
В силу равновероятного расхождения хромосом при формировании гамет и их встречи в зиготе следует ожидать, что в потомстве большого числа браков, подобных только что рассмотренному, у половины сыновей разовьется гемофилия. В то же время все дочери в любом случае получают Х-хромосому от своего отца с геном Х H , поэтому у них всегда нормальная свертываемость крови, но половина дочерей будут гетерозиготными носительницами этого заболевания.
Ген, вызывающий дальтонизм (неспособность различать красный и зеленый цвет), также сцеплен с Х-хромосомой.
Взаимодействие генов
В примерах, которые мы до сих пор рассматривали, каждый ген определял только один признак, и развитие каждого признака зависело только от одного гена. Однако, как правило, отношения между генами и признаками гораздо сложнее.
Множественное действие гена . Один и тот же ген может влиять на формирование ряда признаков организма. Например, ген, вызывающий образование бурой семенной кожуры у гороха, способствует развитию пигмента и в других частях растения. Растения с семенами, покрытыми бурой семенной кожурой, имеют цветки фиолетовой окраски, а растения с белой кожурой семян – белые цветки. Влияние одного гена на развитие многих признаков называют множественным действием гена. Такое множественное действие характерно для большинства генов. Обычно судят о действии гена только по тем признакам, которые легко обнаруживаются. Например, у мухи дрозофилы глаза имеют красную окраску, развитие которой обусловлено присутствием гена W. При наличии аллелей w пигмент, влияющий на окраску глаз, отсутствует, и они становятся белыми. Оказалось, что аллель w влияет также на осветление окраски тела и изменение формы некоторых внутренних органов мухи.
В некоторых случаях проявление множественного признака легко наблюдать. Например, у человека есть ген, определяющий рыжую окраску волос. Этот же ген обусловливает более светлую окраску кожи, а также появление веснушек. У растений гороха ген, отвечающий за красную окраску цветов, определяет и красноватый оттенок стеблей.
Отношение ген – признак . При скрещивании в результате взаимодействия двух различных генов в потомстве могут возникать новые признаки, отсутствовавшие у родительских организмов. Это явление называется новообразованием при скрещиваниях. Его часто наблюдают при разведении домашних животных и культурных растений.
Приведем пример. У душистого горошка – садового растения – есть много сортов, которые отличаются по окраске цветков, в частности, например как в сорте с белыми цветками. При некоторых комбинациях скрещивания двух белых сортов между собой полученные семена дают растения с фиолетовыми цветками. Биохимический анализ показал, что фиолетовая окраска цветков душистого горошка – результат реакции между двумя веществами, каждое из этих веществ образуется под действием определенного гена. Например, есть два гена, каждый с двумя аллелями – С, с и Р, р. Окраска цветков образуется тогда, когда в генотипе растений присутствуют одновременно два доминантных гена С и Р. Скрещивание двух сортов душистого горошка с белыми цветками, когда гибриды первого поколения имеют фиолетовые цветки, записывают следующим образом:
Известно много примеров, когда гены влияют на характер проявления определенного неаллельного гена или на саму возможность проявления этого гена. Пример взаимодействия двух пар генов – наследование формы гребня у кур некоторых пород. В результате различных комбинаций этих генов возникают четыре варианта формы гребня.
Рис. 34. Форма гребня у петухов (А – простой (ааbb), Б – гороховидный (ааВВ или ааВb), В – ореховидный (ААВВ или АаВb), Г – розовидный (ААbb или Ааbb))
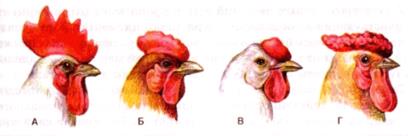
Таким образом, мы видим, что в генотипе любого организма гены взаимодействуют между собой; на фенотипическое проявление признака влияет целый комплекс генов. Сочетания генов в организме обусловливают индивидуальные различия особей одного вида.
Пол как биологический признак. Первичные и вторичные половые признаки.
Пол – это совокупность морфологических, физиологических, биохимических, поведенческих и других признаков организма, обуславливающих репродукцию.
Признаки, по которым отличаются особи разных полов, делят на первичные и вторичные. Первичные представлены органами, обеспечивающими образование гамет и оплодотворение (гонады, половые пути, наружные и внутренние половые органы, которые закладываются в эмбриогенезе). Вторичные – не принимают непосредственного участия в репродукции, развиваются под воздействием половых гормонов и формируются в период полового созревания. Это морфологические признаки организма. Например, особенности развития костно-мышечной системы, подкожной жировой клетчатки, волосяного покрова, тембра голоса, поведения животных.
Признаки особи, связанные с полом, можно разделить на 3 категории:
Ограниченные полом;
Контролируемые полом;
Сцепленные с полом.
Развитие первых обусловлено генами, расположенными в аутосомах обоих полов, но проявляющихся только у одного пола. Такое явление наблюдается в связи с воздействием соответствующих половых гормонов.
Примером вторых является разная степень проявления генов у лиц разного пола. Так у человека ген лысости – только у мужчин, а подагра – у 40% мужчин и редко – у женщин.
Хромосомная и балансовая теории определения пола.
Типы определения пола:
Прогамный – до оплодотворения, по строению мужских и женских гамет.
Сингамный – генетическое определение пола при оплодотворении, которое зависит от характера сочетания половых хромосом, либо от соотношения половых хромосом и аутосом.
Эпигамный – формируется под влиянием внешней среды.
К сингамному типу относится хромосомное определение пола. Ответственные за пол хромосомы назвали половыми. Нормальная мужская гамета несет либо Х либо Y -хромосому, а все яйцеклетки – Х -хромосому. В случае нормального расхождения хромосом при мейозе образуются нормальные яйцеклетки и сперматозоиды с обычным набором хромосом Х или Y . Пол зиготы определяется по соотношению хромосом в гаметах. При этом различают гомогаметный и гетерогаметный пол. У гомогаметного пола одинаковые гаметы. Например, у млекопитающих, дрозофилы гомогаметный женский пол – ХХ . У птиц, рептилий, насекомых (бабочки) гомогаметным является мужской пол ZZ .
Хромосомная теория пола К. Корренса (1907) заключается в том, что пол определяется сочетанием половых хромосом при оплодотворении. Различают следующие типы хромосомного определения пола: ХY, Х0, ZW, Z0 (Табл. 5).
Таблица 5 - Хромосомное определение пола
|
Типы хромосомного определения пола |
Сочетание половых хромосом |
типы гамет |
||
|
Гетерогаметность мужского пола |
||||
|
Прямокрылые насекомые (клопы Protenor, жуки, пауки, кузнечики) | ||||
|
Дрозофилы, позвоночные (млекопитающие, в том числе человек) | ||||
|
Гетерогаметность женского пола |
||||
|
Птицы, рыбы, бабочки, шелкопряд, рептилии, земноводные | ||||
|
Моли и другие беспозвоночные | ||||
При нарушении течения митоза или мейоза могут образовываться особи-гинандоморфы. Содержание половых хромосом в разных клетках таких особей может быть разное (мозаичное). Случаи мозаицизма: ХХ/ХХХ, XY/XXX; X0/XXY и др.
При нерасхождении половых хромосом в гаметогенезе возможны их комбинации, что является причиной хромосомных аббераций у человека (Табл. 6).
В случае нерасхождения половых хромосом при мейозе образуются гаметы ХХ и 0 у самок, а так же ХY и 0 – у самцов. При участии их в оплодотворении формируются зиготы с необычным сочетанием половых хромосом. У человека такие аномалии встречаются 1 на 600-700 новорожденных. Зигота Y0 погибает на ранней стадии; особи ХХХ, ХХY, Х0 – жизнеспособны. Избыток Х -хромосом вызывает конституциональные аномалии и дефекты интеллекта.
Таблица 6 - Возможные комбинации половых хромосом у человека
Балансовая теория пола (К.Б. Бриджес, 1922) подразумевает, что в определении пола принимают участие не только половые хромосомы, но и аутосомы. Один гаплоидный набор аутосом сообщает особи свойства мужского пола. В данном случае пол определяется соотношением количества половых хромосом к набору аутосом.
Гены женского организма сосредоточены в Х -хромосомах, мужского – в аутосомах (А ).
самки имеют баланс 2Х: 2А=1
самцы – 1Х: 2А=0,5.
Нормальный баланс половых хромосом и аутосом у человека:
женщин – ХХ: 44А. (2х: 2А)
мужчин – ХY: 44А. (1х: 2А)
Нарушения:
ХО: 44А. – моносомия у женщин.
ХХХ: 44А. – трисомия у женщин.
ХХY, ХХХY: 44А. – синдром Кляйнфельтера (мужской фенотип)
ХYY: 44А. – полисомия по Y.
Определение, дифференцировка и переопределение пола в онтогенезе.
Формирование половых признаков осуществляется под генетическим контролем. Генетический пол зародыша человека определяется набором половых хромосом при слиянии гамет: ХХ и Х Y .
Зачатки гонад у ранних эмбрионов до 5-й или 6-й недели не различаются у разных полов и называются бисексуальными. Они состоят из внешнего слоя – кортекса (cortex) и внутреннего слоя – медулла (medulla). Первичные клетки зародышевого пути обнаруживаются у человека на 3-й неделе эмбрионального развития в эндодерме желточного мешка. Затем под влиянием хемотаксических сигналов они мигрируют в гонады. Эта миграция не зависит от пола. Зачатки гонад могут развиваться в яичники или семенники. Дифференцировка мужской гонады наблюдается на 7-й неделе. На 36 день семенник начинает выделять андрогены (тестостерон), определяющий развитие мужского пола. Развитие женской гонады наблюдается на 8-й неделе. Результатом этого является образование женских гормонов – эстрогенов.
В норме Х -хромосомы содержат ген-репрессор – ген тестикулярной феминизации Tfm. Нормальная аллель гена Tfm определяет синтез белкового рецептора для андрогенов. Эти рецепторы имеются на поверхности клеток гонад обоих типов. Развитие по мужскому фенотипу зависит от гена Y -хромосомы – H- y -антигена . Его секретируют первичные мужские клетки зародышевого пути. H-y-антиген отвечает за выработку тестостерона. Как только эти клетки попадают в зачатки гонад, начинается дифференцировка семенников. Считалось, что мужской фенотип определяется всей мужской Y -хромосомой. Но в 1990 году был открыт ген (Sex Region Y ), локализованный в Y -хромосоме. При его отсутствии генотип XY дает женский фенотип.
При сочетании половых хромосом ХY , белки-рецепторы воспринимают андрогены. Формируется нормальный мужской фенотип. В случае рецессивной мутации (t fm ) рецепторы на поверхности клеток не синтезируются и гормон не воспринимается. Возникает несоответствие между мужским генотипом Х t fm Y и формирующимся женским фенотипом (синдром Морриса).
Зачаточная гонада у человека бисексуальна до 6-й недели внутриутробного развития (Табл. 7).
Генотип особи XY вызывает развитие медуллы и формирует семенник на 7-8-й неделе.
Генотип особи ХХ вызывает развитие кортекса и формирует яичник на 8-9-й неделе внутриутробного развития.
Гонады детерминируют развитие первичных и вторичных половых признаков. Половые железы выделяют гормоны, которые вместе с гормонами эндокринных желез контролируют пути дифференцировки пола. Уровень гормонов в свою очередь контролируется генами.
Таким образом, процесс половой дифференцировки включает:
Генетический контроль;
Регуляторные функции гормонов.
Существует теория действия гормонов в качестве регуляторных факторов на гены. Они действуют только на специфические клетки-мишени . В клетке вырабатывается особый белок – рецептор, связывающийся с гормоном по заданному типу развития. После чего гормон приобретает свойства индуцировать работу одного или нескольких генов в хромосомах. Клетки-мишени женского организма воспринимают гормоны по женскому типу в большей степени, а клетки-мишени мужского организма – мужские гормоны. Соответственно в норме формируется женский или мужской фенотип. Таким образом, существует следующая схема:
В клетках-мишенях вырабатывается белок-рецептор;
Белок-рецептор связывается с гормоном;
Инициируется работа нескольких генов в хромосомах.
Образование белков-рецепторов и гормонов контролируется в свою очередь генами. В случае мутаций возникает нарушение контроля, вызывающие аномалии. Примером служит описанный выше синдром Морриса или тестикулярной феминизации. У лиц с этим заболеванием отсутствуют белки-рецепторы к тестостерону и гормон не воспринимается. В силу этого, развитие по мужскому типу прекращается, и появляются женские фенотипические признаки. В исключительных случаях возможно исправление таких дефектов введением соответствующих гормонов.
Мутации таких генов вызывают:
Нарушения синтеза белков-рецепторов;
Отсутствие восприятия гормонов;
Нарушение формирования пола.
Нормальная аллель, контролирующая синтез белкового рецептора для андрогенов – Х Tfm . Рецессивная мутация гена, вызывающая нарушение синтеза рецептора – Х tfm . . При этом гормон по заданному типу не воспринимается.
Особенности детерминации пола у человека.
Таблица 7 - Этапы дифференцировки пола у человека
|
Периоды внутриутробного развития (недели). |
Развитие гонад и половых признаков. |
|
первичные половые клетки |
|
|
закладка первичных гонад |
|
|
гонада бисексуальна |
|
|
развитие гонады по мужскому типу (семенников) |
|
|
развитие гонады по женскому типу (яичников) |
|
|
формирование гормонального пола |
|
|
формирование внутренних половых органов |
|
|
формирование наружных половых органов |
Таблица 8 - Половой диморфизм
|
Генетический: |
сочетание хромосом: ХХ или ХY . |
|
Гаметный: |
наличие гамет: яйцеклеток, сперматозоидов. |
|
Гонадный: |
формирование желез: семенников или яичников. |
|
Гормональный: |
функция мужских гормонов – тестостерона, женских гормонов – прогестерона, этрогена. |
|
Фенотипический: |
формирование вторичных половых признаков по мужскому или женскому типу. |
|
Психологический: |
половая и поведенческая принадлежность. |
Нарушение полового самосознания.
Изначальная генетическая бисексуальность гонад является основой переопределения пола.
В результате нарушения воздействия гормонов или функций рецепторов клеток-мишеней может происходить переопределение пола.
В природе много факторов, ослабляющих действие генов, которые контролируют развитие пола. Например, у человека в гонадах одной особи могут развиваться в равной степени семенниковая и яичниковая часть.
Гермафродитизм – явление интерсексуальности.
На основании клинических данных различают 3 типа интерсексов:
Истинный гермафродитизм – наличие у особи гонад и половых клеток обоих полов;
Мужской псевдогермафродитизм: имеются только тестикулы (семенники), фенотип – женский;
Женский псевдогермафродитизм: имеются только яичники, фенотип – мужской.
Соотношение полов:
Первичное – в момент оплодотворения соотношение должно быть близким 1:1, так как встреча половых клеток равновероятна. При обследовании у человека обнаружено, что на 100 женских зигот образуется 140‑160 мужских. Сперматозоиды, содержащие Y-хромосому, легче, подвижнее и имеют большой отрицательный заряд, в то время как яйцеклетка имеет положительный заряд. Поэтому, Y-содержащие сперматозоиды чаще оплодотворяют яйцеклетку.
Вторичное – к моменту рождения на 100 девочек рождается 103-105 мальчиков. К 20 годам на 100 девушек приходится 100 юношей.
Третичное – к 50 годам на 100 женщин приходится 85 мужчин, а к 85 годам на 100 женщин – 50 мужчин. Считается, что женский организм более приспособленный, что может быть объяснено наряду с другими причинами мозаицизмом женского организма по половым хромосомам.
В 1962 году М. Лайон высказала гипотезу об инактивации одной Х‑хромосомы у женского организма млекопитающих. У женского зародыша функционируют обе хромосомы до 16 дня эмбрионального развития. На 16-й день происходит инактивация одной хромосомы с образованием полового хроматина (тельца Барра). Процесс этот случайный, поэтому примерно в 1/2 клеток активной сохраняется материнская Х-хромосома, а отцовская инактивируется. В других – отцовская активна, а материнская хромосома инактивируется. Переактивация не происходит. Материнская и отцовская Х-хромосомы содержат аллельные, но не абсолютно одинаковые гены, т.е. в одной хромосоме может быть локализован доминантный аллель, а в другой – рецессивный. Обладание дополнительными аллелями расширяет приспособительные возможности организма.
Женский организм более устойчив к холоду, ионизирующему излучению, эмоциональным перегрузкам (женщины плачут чаще, вместе со слезами выделяются активные амины, в результате снижается кровяное давление).
Однако, если бы гипотеза работала без ограничений, то не было бы фенотипических различий между здоровыми женщинами с двумя Х‑хромосомами и больными Х0, или у мужчин XY и XXYY. Очевидно, Х‑хромосома инактивируется не полностью.
У растений и животных хромосомный механизм определения пола является наиболее распространённым. Согласно хромосомной теории, пол организма определяется половыми хромосомами (гоносомами). В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомной детерминации:
· самки гомогаметны, самцы гетерогаметны (Drosophila-тип)
o самки XX самцы XY (Lygaeus-тип)
o самки XX самцы X0 (Protenor-тип)
· самки гетерогаметны, самцы гомогаметны (Abraxas-тип)
o самки ZW самцы ZZ
o самки Z0 самцы ZZ
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковые половые хромосомы, которые обозначаются как XX (ZZ ). Организмы такого пола продуцируют гаметы только одного класса- содержащие по одной X (Z ) хромосоме.
У особей же гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W ), либо только одна- X (Z ) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом.
У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, некоторые рыбы, растения и др.
Существует два основных типа XY-детерминации пола:
· Как у человека: пол зависит от наличия Y -хромосомы (если она есть, проявляется фенотип самца, если нет - самки). Так, при мутациях, связанных с изменением числа половых хромосом, особи, имеющие в генотипе комбинации XY , XXY , XYY , XXYY и т. п. будут обладать мужским фенотипом различной степени выраженности, а имеющие комбинации XX , X , XXX , XXXX и т. п. - женским.
· Как у представителей рода мушек-дрозофил: пол определяется по соотношению числа X -хромосом и числа аутосом, наличие же Y -хромосомы никак не влияет на детерминацию пола, но самцы без неё стерильны, так как в ней находятся гены, ответственные за сперматогенез.
У многих других организмов (птицы, некоторые рептилии и рыбы, бабочки, ручейники, из растений - земляника) наблюдается обратная картина - гомогаметен мужской пол (имеет две гомологичные хромосомы Z ), а гетерогаметен женский (имеет одну Z -хромосому и одну состоящую в основном из гетерохроматина и потому генетически инертную W -хромосому).
Вероятно, исходным для бабочек механизмом определения пола был механизм ZO самка/ZZ самец. Затем, путем хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У видов с системой Z/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом, но у тутового шелкопряда (система WZ/ZZ) обнаружен отвечаюший за развитие женского пола ген Fem в W-хромосоме.
При этом механизме определения пола один из полов (гомогаметный) обладает двумя X-хромосомами, в то время, как второй (гетерогаметный) только одной. При этом пол определяется так же, как и у дрозофил: по соотношению числа X-хромосом и аутосом. Этот механизм определения пола обнаружен у некоторых насекомых (клопов, бабочек и др.) и круглых червей. У нематоды Caenorhabditis elegans при наборе половых хромосом XX формируется гермафродит, а при наборе ХО - самец.
Гетерохромосомы.
Хромосомы, отличающиеся некоторыми особенностями во время деления (син. аллосомы); в последнее время часто применяется в более узком смысле: хромосомы, имеющиеся в соматических клетках особей одного из полов (в большинстве случаев мужского) в одиночном числе (син. идио-хромосомы, половые хромосомы), в противоположность аутосомам, к-рых имеется всегда по паре каждого типа. Т. о., первоначальное понятие включает в себя гетерохро-мосомы в смысле современных авторов, а также нек-рые другие типы хромосом, например, мелкие хромосомы у нек-рых Rhynchota (клопов). В одних случаях в клетках самцов имеется одна непарная хромосома и соответственно ей в клетках самок 2 хромосомы того же типа. Такие хромосомы называются ж-хромосомами. Кроме ж-хромосомы, у самцов бывает часто еще одна непарная хромосома-г/-хромосома – Более редко (бабочки, птицы)-у самок одна ж-хромосома или ж – и ^/-хромосомы, а у самцов 2 ж-хромосомы. Нек-рые авторы для этого случая дают особые названия: вместо ж-хромосомы-г-хромосома, вместо у -w. Иногда имеется не одна ж-хромосома, а несколько (2, 5, 6 и более); в таком случае разница между обоими полами по числу хромосом сильно увеличивается. Пол, имеющий одну ж-хромосому (или по одной ж-хромосоме каждого типа), называется гетерозиготным или гетерогаметным, с двойным количеством ж-хро-мосом - гомозиготным или гомогаметным. Половые клетки гетерозиготного пола после редукционного деления будут двух типов: с ж-хромосомой и без нее (и с ^-хромосомой, если она имеется); у гомозиготного пола все зрелые половые клетки одинаковы, с ж-хромосомой. Открытие гетерохромосом имело решающее значение в выяснении вопросов определения пола (см.) и связанного с полом наследования. В виду отношения Г. к определению пола их обычно называют половыми хромосомами. Другой синоним- идиохромосомы-сначала применялся только для типа ху; такого обозначения придерживаются и до сих пор некоторые авторы, называя гетерохромосомами только ж-хромосомы в случае отсутствия.
Пол организмов, размножающихся половым способом, определяется парой особых половых хромосом, тогда как остальные хромосомы называют аутосомами.
Пол, особи которого имеют одинаковые половые хромосомы, называется гомогаметным. У особей гомогаметного пола при образовании гамет все они будут одинаковыми по половым хромосомам.
Пол, у особей которого половые хромосомы разные, называется гетерогаметным. Соответственно и половые клетки этих особей будут двух типов, причем образовываться они будут в равном соотношении.
У половых хромосом нет принципиальных отличий от аутосом: они, как и аутосомы, несут гены, которые контролируют развитие тех или иных признаков.
Половые хромосомы принято обозначать буквами. У видов с гетерогаметным мужским полом одинаковые по размеру и форме половые хромосомы обозначают буквой X, а хромосому, имеющуюся только у самцов, - буквой Y.
Такой тип наследования пола встречается у многих червей, моллюсков и членистоногих, а из позвоночных - у млекопитающих и человека. В хромосомном комплексе самок этих видов две X-хромосомы, а у самцов - X-хромосома и Y-хромосома.
У видов с гетерогаметным женским полом хромосому, имеющуюся как у самок, так и у самцов обозначают буквой Z, а имеющуюся только у самок - буквой W. Самки этих видов имеют хромосомный комплекс с половыми хромосомами ZW, а самцы - с половыми хромосомами ZZ. Такой тип наследования пола имеет место, в частности, среди растений - у клубники, среди насекомых - у бабочек, а среди позвоночных - у рыб, пресмыкающихся и птиц.
Медоносная плела относится к видам, у которых гомогаметным является женский пол, но пчелиная матка откладывает два типа яиц - оплодотворенные и неоплодотворенные. Из оплодотворенных яйцеклеток (генотип XX) развиваются самки (матка и рабочие пчелы), а из неоплодотворенных (одна X- хромосома, генотип X0) - самцы (трутни). В связи с этим организмы, образовавшиеся при обыкновенном половом размножении, - диплоидные, а самцы, развивающиеся в результате партеногенеза, - гаплоидные. В процессе индивидуального развития в клетках тела трутней восстанавливается диплоидный хромосомный набор, тогда как в половых клетках гаплоидное число хромосом сохраняется и при образовании сперматозоидов не уменьшается. Аналогичное явление встречается и у некоторых других насекомых.
Х- хромосома намного длиннее, чем Y-хромосома, и содержит некоторые гены, которых нет в Y-хромосоме, в том числе и гены, кодирующие не связанные с проявлением пола белки.
♀XX, ♂X0 (пустынная саранча)
♀XX, ♂XY(дрозофила, млекопитающие)
♀XY, ♂XX(бабочки, птицы, рептилии)
♀X0, ♂XX(тля, моль)
♀2n, ♂nгапло-диплоидный тип (пчелы)
На базе генетической информации в зависимости от второй хромосомы на 2-12 неделях эмбриогенеза развивается гонадный пол – яичники или семенники. Они выделяют соответственно эстрогены или андрогены, под действием которых формируется гормональный пол. Одновременно определяется морфо-физиологический пол (мужской или женский фенотип)