40. Гомо и гетерогаметный пол
В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для нормального самовоспроизведения вида. Детерминация пола может происходить на разных этапах размножения. Различают три основных типа детерминации:
1) эпигамный, когда пол особи определяется в процессе онтогенеза. Детерминация пола в данном случае значительно зависит от внешней среды;
2) прогонный, когда пол будущего дочернего организма определяется в ходе гаметогенеза у родителей особи;
3) сингамный, при котором пол дочерней особи определяется в момент слияния гамет. Это наиболее распространенный тип детерминации пола, характерный для животных всех видов.
При прогамном и сингамном типах детерминации пол зависит от определенных половых хромосом.
У самок млекопитающих в диплоидном наборе хромосом выделяют пару одинаковых по форме гоносом (половых хромосом), обозначаемых ХХ-хромосомами. Самцы в кариотипе содержат Х- и Y-хромосомы. Самки птиц содержат две разные (XY), а самцы одинаковые (XX) половые хромосомы. В период редукционного деления (мейоза) у самок млекопитающих образуется один тип гамет с Х-хромосомой, поэтому женский пол называют гамогаметным. У самцов образуется два типа гамет с Х- и Y-хромосомами, поэтому мужской пол называют гетерогаметным. Определение пола млекопитающих зависит от того, каким спермием будет оплодотворена яйцеклетка. Если яйцеклетка оплодотворена спермием, содержащим Х-хромосому, то происходит закладка особи женского пола, если спермий несет Y-хромосому - закладывается особь мужского пола.
Схема образования гамет в мейозе самцов и самок, определения пола при оплодотворении следующая:
Существует равная вероятность закладки особи мужского или женского пола, так как число сперматозоидов у самцов (яйцеклеток у птиц) с Х- и Y-хромосомами одинаково.
Самцов обычно рождается на несколько процентов больше, чем самок, но в ходе эволюции выработался механизм сохранения числового соотношения полов, поэтому к возрасту половой и физиологической зрелости это соотношение выравнивается вследствие более высокой смертности самцов.
43. Опыты п регулированию соотношению пролов
44. Наследование признаков огран. Полом
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, ОГРАНИЧЕННЫХ ПОЛОМ
Наследование признаков, контролируемых генами, локализованными в аутосомах, но фенотипически проявляющихся исключительно или преимущественно у одного пола, - есть наследование, ограниченное полом. Расщепление по таким признакам соответствует правилам Менделя. Выделяют также признаки, контролируемые полом. Они обусловлены генами, локализованными в половых хромосомах, но проявляются у обоих полов по-разному. К числу патологических, или нежелательных, признаков, ограниченных полом, у животных можно отнести гипоплазию вымени и сосков, гипоплазию гонад (семенников и яичников), крипторхизм, недоразвитие разных частей половых органов у телок (наблюдаемое преимущественно у животных белой масти), сужения прямой кишки и матки у коров, аплазию сегментных протоков Вольфа, затянувшуюся беременность у самок, аномалии сперматозоидов у самцов и др. Большинство из указанных аномалий вызывает частичное или полное нарушение воспроизводительной функции.
ПРОБЛЕМА РЕГУЛЯЦИИ ПОЛА
Проблема регуляции пола вытекает из необходимости увеличения продукции животноводства за счет преимущественного получения особей одного вида, дающих более высокий выход молока, мяса, шерсти, яиц и т. д. Так, в молочном скотоводстве более желательно рождение телочек, а в мясном - бычков, так как они быстрее растут. От высокоценных племенных быков и коров целесообразно получать мужских потомков для более быстрого размножения их генотипов. В яичном птицеводстве экономически более выгодно получение курочек. В связи с этими практическими потребностями исследователи не только стремятся познать механизмы определения пола, но и изучают возможности искусственного регулирования пола. Необходимо отметить, что в отношении крупных животных с внутриутробным развитием плодов эта проблема еще не решена. Регуляция соотношения полов у млекопитающих может быть достигнута путем разделения спермы на две фракции: первую - содержащую в спермиях Х-хромосому и вторую - содержащую Y-хромосому. Оплодотворение самок одной из этих фракций будет давать приплод одного пола. Проводились эксперименты по разделению спермы на указанные фракции центрифугированием, электрофорезом и седиментацией (осаждением) с помощью аминокислого гистидина. Осеменение самок крольчих, например, более легкой и более подвижной фракцией приводило к сдвигу в сторону мужского пола. Однако полного сдвига в соотношении полов сделано не было. Разрабатывается метод количественного определения ДНК в спермиях путем измерения интенсивности флуоресценции ядер. Полученные результаты, как считают авторы этого метода, могут стать предпосылкой для успешного разделения спермиев у млекопитающих на несущие X-или Y-хромосому.
Партеногенез. Это развитие организма без оплодотворения. Получение особей одного пола может быть достигнуто при развитии эмбрионов из отцовских (андрогенез) или материнских (гиногенез) гамет. Так, под руководством Б. Л. Астаурова были проведены эксперименты по андрогенезу у тутового шелкопряда. Неоплодотворенные яйца шелкопряда подвергали тепловому шоку и облучали рентгеном, тем самым разрушали их ядра, не повредив цитоплазму. Затем эти яйца осеменяли. Зигота формировалась путем слияния ядер двух проникших в яйцо спермиев, развившиеся из нее особи имели признаки только отцовского вида.
В другом варианте экспериментов неоплодотворенные и еще не прошедшие редукционного деления яйца нагревали, останавливая тем самым мейоз и сохраняя диплоидность набора хромосом. Из таких яиц без оплодотворения (партеногенетически) развивались только самки, унаследовавшие признаки матери. Девственное развитие (партеногенез) в естественных или спонтанных условиях встречается у птиц. И. В. Кудрявцев, 3. А. Ощепкова, А. К. Голубев и др. на основании экспериментов, проведенных на курах разных пород, пришли к выводу, что существуют генетические предпосылки селекции на получение жизнеспособных особей - партеногенов и создание линий с высокой предрасположенностью к партеногенезу. При этом все вылупляющиеся цыплята оказываются петушками. Отбор на повышение частоты партеногенеза, проведенный в двух линиях индеек, дал следующие результаты. Способность яиц к партеногенезу возросла с 1,1 до 18,6 % в первой линии ис4до21,1%во второй. Значение партеногенеза не только в том, что он позволяет получить потомство одного пола, но и в том, что использование этого явления дает возможность получить особей, идентичных генотипу одной из родительских форм.
Подавляющее большинство животных представлено особями двух полов - мужского и женского. У некоторых видев растений также наблюдаются особи двух полов (конопля, печеночный мох сферокарпус, дрема и др.). Механизм, определяющий пол особи, стал ясен после создания хромосомной теории наследственности, хотя некоторые наблюдения о закономерностях наследования пола были сделаны намного раньше. Давно известно, что в среднем в популяции вида соотношение особей мужского и женского пола одинаково, т. е. равно 1:1. Такое соотношение у Г. Менделя вызвало аналогию с моногибридным анализирующим скрещиванием, как описано выше, при анализирующем скрещивании расщепление 1:1 возникает в случае, если один из родителей гетерозиготен по изучаемому гену (Аа), а другой - гомозиготен по рецессивному (аа). Соотношение 1:1, наблюдаемое при расщеплении по полу, давало возможность предположить, что один из полов гетерозиготен, а другой - гомозиготен по фактору, определяющему пол организма. Это предположение еще в начале века было подтверждено результатами генетических экспериментов и получило цитологическое подтверждение после открытия половых хромосом. При изучении строения хромосом половых клеток животных и клеток их тела было установлено, что самцы и самки различаются по набору хромосом. Детально половые хромосомы были изучены У. Вильсоном и К. Мак Клангом.
Половые хромосомы. Современное представление о хромосомном механизме определения пола удобно рассмотреть на хромосомах дрозофилы. На рис. 84 схематически представлен хромосомный набор дрозофилы, состоящий всего из восьми хромосом, составляющих 4 пары. В трех парах хромосомы гомологичны друг другу и морфологически не отличаются. Хромосомы четвертой пары сильно различаются. Одна из них, называемая Х-хромосомой,- палочковидная, вторая (У-хромосома) имеет форму крючка, образованного неравными по величине плечами этой хромосомы, В клетках самок четвертая пара у дрозофилы представлена двумя Х-хромосомами, у самцов
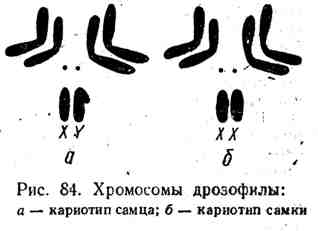
Одной X и одной У-хромосомой. Пара хромосом, по которой различаются самцы и самки, называется половыми хромосомами или гетерохромосомами в противоположность остальным хромосомам, именуемым аутосомами (иногда они сокращенно обозначаются А).
Гомо- и гетерогаметный пол. У самок дрозофилы, имеющих одинаковые половые хромосомы (XX ), все возникшие в результате мейоза половые клетки имеют одинаковую половую хромосому (X ), вследствие чего все гаметы одинаковы. Женский пол дрозофилы, продуцирующий гаметы только одного типа, называется гомогаметным. В свою очередь самцы дрозофилы продуцируют два сорта гамет, в одном из которых половая хромосома представлена Х-хромосомой, а в другом - У-хромосомой. Мужской пол у дрозофилы является в силу этого гетерогаметным.
В зависимости от принадлежности самок к гомо- или гетерогаметному полу всё животные делятся на две группы. У млекопитающих, червей, ракообразных, большинства насекомых (в том числе дрозофилы), большинства земноводных, некоторых рыб гетерогаметным является мужской пол, гомогаметным - женский.
К этой группе относится и человек. В соматических клетках человека содержится по 44 аутосомных хромосомы и, кроме того, у женщин две Х-хромосомы, а у мужчин - одна Х-хромосома и одна У-хромосома. Хромосомная формула женского организма человека может быть обозначена 44А + XX , мужского - 44А + ХУ. У птиц, пресмыкающихся, некоторых земноводных и рыб, части насекомых (бабочки и ручейники) гетерогаметным является женский пол. В этом случае для обозначения половых хромосом используют другие символы. При гетерогаметности женского пола его половые хромосомы обозначают ZW , а половые хромосомы мужского пола, - ZZ . Например, у кур, имеющих в соматических клетках 78 хромосом, хромосомная формула мужского пола -76А + ZZ , женского - 76А+ ZW .
У некоторых видов в процессе эволюции произошла утрата одной из половых хромосом гетерогаметного пола, в результате чего в их хромосомном наборе присутствует лишь одна половая хромосома. Формула половых хромосом в таком случае у гетерогаметного пола обозначается как ХО или ZO .
Механизм расщепления по полу 1:1. Знание хромосомного механизма определения пола позволяет объяснить причины появления мужских и женских особей в соотношении 1:1. Как отмечалось выше, гомогаметный пол образует гаметы одного типа, каждая из которых несет одинаковую половую хромосому. В свою очередь в гаметах гетерогаметного пола представлена либо одна, либо другая половая хромосома. Например, у дрозофилы все половые клетки самок имеют одну Х-хромосому, половина гамет самца имеет Х-хромосому, другая половина - У-хромосому.
Если яйцо ( X ) оплодотворяется спермием с Х-хромосомой, то сочетание в зиготе двух Х-хромосом ведет к образованию самки (XX ). Оплодотворение того же яйца спермием с У-хромосомой обусловливает появление самца (ХУ). В связи с тем что участие в оплодотворении того или другого спермия равновероятно, возникает равная вероятность появления самцов и самок. Учет пола у большого числа новорожденных особей у животных разных видов показывает соотношение 1:1 или очень близкое к нему. Так, из каждых 100 новорожденных особи мужского пола составляют у человека 51, у собак - 56, у кур - 49, у мышей - 50, у овец - 49, у крупного рогатого скота-51, у лошадей - 52 Приведенные цифры характеризуют соотношение особей разного пола в момент рождения. В дальнейшем эти соотношения могут сильно изменяться в силу неодинаковой выживаемости особей разного пола. Так у человека к 50 годам соотношение мужчин и женщин составляет 85:100, а к 85 годам - 50: 100.
Обусловленное хромосомным механизмом проявление того или иного пола в ряде случаев может быть экспериментально изменено, что показано на курах, тутовом шелкопряде и др. У двудомных растений, имеющих мужские и женские растения, также изучены половые хромосомы; гетерогаметным у большинства таких растений является мужской пол.
Большинство животных являются раздельнополыми организмами. Пол можно рассматривать как совокупность признаков и структур, обеспечивающих способ воспроизводства потомства и передачу наследственной информации. Пол чаще всего определяется в момент оплодотворения, то есть в определении пола главную роль играет кариотип зиготы. Кариотип каждого организма содержит хромосомы, одинаковые у обоих полов, - аутосомы, и хромосомы, по которым женский и мужской пол отличаются друг от друга, - половые хромосомы. У человека «женскими» половыми хромосомами являются две Х-хромосомы. При образовании гамет каждая яйцеклетка получает одну из Х-хромосом. Пол, у которого образуются гаметы одного типа, несущие Х-хромосому, называется гомогаметным. У человека женский пол является гомогаметным. «Мужские» половые хромосомы у человека - Х-хромосома и Y-хромосома. При образовании гамет половина сперматозоидов получает Х-хромосому, другая половина - Y-хромосому. Пол, у которого образуются гаметы разного типа, называется гетерогаметным. У человека мужской пол - гетерогаметный. Если образуется зигота, несущая две Х-хромосомы, то из нее будет формироваться женский организм, если Х-хромосому и Y-хромосому - мужской.
У животных можно выделить следующие четыре типа хромосомного определения пола .
1. Женский пол - гомогаметен (ХХ), мужской - гетерогаметен (ХY) (млекопитающие, в частности, человек, дрозофила).
Генетическая схема хромосомного определения пола у человека:
Генетическая схема хромосомного определения пола у дрозофилы:
2. Женский пол - гомогаметен (ХХ), мужской - гетерогаметен (Х0) (прямокрылые).
Генетическая схема хромосомного определения пола у пустынной саранчи:
3. Женский пол - гетерогаметен (ХY), мужской - гомогаметен (ХХ) (птицы, пресмыкающиеся).
Генетическая схема хромосомного определения пола у голубя:
Яндекс.ДиректВсе объявления
4. Женский пол - гетерогаметен (Х0), мужской - гомогаметен (ХХ) (некоторые виды насекомых).
Генетика пола
Пол характеризуется комплексом признаков, определяемых генами, расположенными в хромосомах. В клетках организма человека хромосомы составляют парные диплоидные наборы. У видов с раздельнополыми особями хромосомный комплекс самцов и самок неодинаков и различается по одной паре хромосом (половые хромосомы). Одинаковые хромосомы этой пары назвали X (икс) -хромосомой, непарную, отсутствующую у другого пола - У (игрек) -хромосомой; остальные, по которым нет различий, - аутосомами (А).
Клетки женщины содержат две одинаковые половые хромосомы, которые обозначаются XX, у мужчин они представлены двумя непарными хромосомами X и Y. Таким образом, набор хромосом мужчины и женщины отличается только одной хромосомой: хромосомный набор женщины содержит 44 аутосомы + XX, мужчины - 44 аутосомы + XY.
Во время деления и созревания половых клеток у человека образуются гаметы с гаплоидным числом хромосом: яйцеклетки, как правило, содержат 22 + Х-хромосомы. Таким образом, у женщин образуется только один тип гамет (гаметы с Х-хромосомой). У мужчин гаметы содержат 22 + X или 22 + Y хромосом, и образуется два типа гамет (гамета с Х-хромосомой и гамета с Y-хромосомой). Если при оплодотворении в яйцеклетку попадает сперматозоид с Х-хромосомой, формируется зародыш женского пола, а с Y-хромосомой - мужского пола.
Следовательно, определение пола человека зависит от наличия в мужских половых клетках - сперматозоидах, оплодотворяющих яйцеклетку, X- или Y-хромосом.
Существует четыре основных типа хромосомного определения пола:
1. Мужской пол гетерогаметен; 50% гамет несут Х-, 50% -У – хромосому например, человек, млекопитающие, двукрылые, жуки, клопы (Слайд 4).
2. Мужской пол гетерогаметен; 50% гамет несут Х-, 50%– не имеют половой хромосомы, например, кузнечики, кенгуру (Слайд 7).
3. Женский пол гетерогаметен; 50% гамет несут Х- , 50% гамет– У– хромосому, например, птицы, пресмыкающиеся, хвостатые амфибии, шелкопряд (Слайд 7).
4. Женский пол гетерогаметен; 50% гамет несут Х- , 50% не имеют половой хромосомы, например, моль.
Наследование признаков, гены которых локализованы в половых хромосомах, называют наследованием, сцепленным с полом.
26. Генотип как целостная система. Взаимодействие генов, множественное действие генов.
Генотип как целостная система
Свойства генов. На основании знакомства с примерами наследования признаков при моно- и дигибридном скрещивании может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов, каждый из которых определяет развитие только своего признака или свойства. Такое представление о прямой и однозначной связи гена с признаком чаще всего не соответствует действительности. На самом деле существует огромное количество признаков и свойств живых организмов, которые определяются двумя и более парами генов, и наоборот, один ген часто контролирует многие признаки. Кроме того, действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводит к заметному изменению генотипа во времени.
Характер проявления действия гена в составе генотипа как системы может изменяться в различных ситуациях и под влиянием различных факторов. В этом можно легко убедится, если рассмотреть свойства генов и особенности их проявления в признаках:
Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов.
Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма.
Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена).
Один ген может влиять на развитие разных признаков - это множественное, или плейотропное, действие гена.
Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков) - это множественные гены, или полигены.
Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков. Такое взаимодействие осуществляется опосредованно - через синтезированные под их контролем продукты своих реакций.
Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды.
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов -неаллельными.
Выделяют следующие основные типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование -тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого. Такое явление возможно при следующих условиях: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака.
Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы (рис. 3.5). Как видно из схемы, гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) - розовые. При скрещивании растения с красными цветками и растения с белыми цветками в F 1 , у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F 2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F 2 по окраске цветков обычно следующее - 1 красная (АА): 2 розовые (Аа): 1 белая (аа).

Рис. 3. 5. Наследование окраски цветков при неполном доминировании у ночной красавицы.
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
Сверхдоминирование - более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса (см. § 3.7).
Кодаминирвание - участие обоих аллелей в определении признака у гетерозиготной особи. Ярким и хорошо изученным примером кодоминирования может служить наследование IV группы крови у человека (группа АВ).
Эритроциты людей этой группы имеют два типа антигенов: антиген А (детерминируемый геном /\ имеющимся в одной из хромосом) и антиген В (детерминируемый геном / а, локализованным в другой гомологичной хромосоме). Только в этом случае проявляют свое действие оба аллеля - 1 А (в гомозиготном состоянии контролирует II группу крови, группу А) и I B (в гомозиготном состоянии контролирует III группу крови, группу В). Аллели 1 А и I B работают в гетерозиготе как бы независимо друг от друга.
Пример наследования групп крови иллюстрирует и прояв-ление множественного аллелизма: ген/может быть представлен тремя разными аллелями, а есть гены, имеющие десятки аллелей. Все аллели одного гена получили название серии мно-жественных аллелей, из которых каждый диплоидный организм может иметь два любых аллеля (и только). Между этими аллелями возможны все перечисленные варианты аллельных взаимодействий.
Явление множественного аллелизма распространено в природе. Известны обширные серии множественных аллелей, определяющих тип совместимости при оплодотворении у грибов, опылении у семенных растений, детерминирующих окраску шерсти животных и т. д.
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого - ааВВ, то
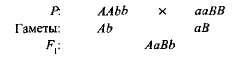
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях {ааВ- и A-bb) цветки у растения белые (знак «-» в формуле генотипа обозначает, что это место может занять как доминантный, так и рецессивный аллель).
При самоопылении растений душистого горошка из F 1 в F 2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/1 6 растений, белые - у 7/16. Решетка Пеннета наглядно показывает причину этого явления (рис. 3.6).
Эпистаз - это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический.
По изменению числа и соотношения фенотип и чес ких классов при дигибридном расщеплении в F 2 рассматривают несколько типов эпистатических взаимодействий: доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>В или b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипичес-ким выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F 1 образуются растения с плодами треугольной формы. При их самоопылении в F 2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково- А 1 и A 2 .

Рис. 3.6 . Наследование окраски цветков у душистого горошка
Тогда все генотипы (А 1 ,-А 2 ,-, А 1 -а 2 а 2 , a 1 a 1 A 2 -) будут иметь одинаковый фенотип - треугольные стручочки, и только растения а 1 а 1 а 2 a 2 будут отличаться -- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А 1 А 2 А 3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А 1 А 1 А 2 А 2 А 3 ,А 3 а зерна а 1 а 1 а 2 a 2 а 3 а 3 не имели пигмента.
По типу кумулятивной полимерии наследуются многие признаки: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Таким образом, многочисленные наблюдения свидетельствуют о том, что проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
Взаимодействие генов
Отношение между генами и признаками достаточно сложное. В организме не всегда один ген определяет только один признак и, наоборот, один признак определяется только одним геном. Чаще один ген может способствовать проявлению сразу нескольких признаков, и наоборот. Генотип организма нельзя рассматривать как простую сумму независимых генов, каждый из которых функционирует вне связи с другими. Фенотипное проявления того или иного признака являются результатом взаимодействия многих генов.
Множественное действие генов (плейотропия) - процессы влияния одного гена на формирование нескольких признаков.
Например, у человека ген, определяющий рыжую окраску волос, обусловливает более светлую кожу и появление веснушек.
Иногда гены, определяющие морфологические признаки, влияют на физиологические функции, снижая жизнестойкость и плодовитость, или оказываются летальными. Так, ген, вызывающий голубую окраску у норки, снижает ее плодовитость. Доминантный ген серой окраски у каракулевых овец в гомозиготном состоянии детален, поскольку у таких ягнят недоразвит желудок и они погибают при переходе на питание травой.
Комплементарное взаимодействие генов. На развитие одного признака могут влиять несколько генов. Взаимодействие нескольких неаллельных генов, приводящее к развитию одного признака, называется комплементарным. Например, у кур имеются четыре формы гребня, проявление какой-либо из них связано со взаимодействием двух пар неаллельных генов. Розовидный гребень обусловлен действием доминантного гена одной аллели, гороховидный - доминантного гена другой аллели. У гибридов при наличии двух доминантных неаллельных генов образуется ореховидный гребень, а при отсутствии всех доминантных генов, т.е. у рецессивной гомозиготы по двум неаллельным генам, образуется простой гребень.
Результатом взаимодействия генов является окраска шерсти у собак, мышей, лошадей, форма тыквы, окраска цветков душистого горошка.
Полимерия - такое взаимодействие неаллельных генов, когда степень развития признака зависит от общего количества доминантных генов. По этому принципу наследуется окраска зерен овса, пшеницы, цвет кожи у человека. Например, у негров в двух парах неаллельных генов 4 доминантных, а у людей с белой кожей - ни одного, все гены рецессивные. Сочетания разного количества доминантных и рецессивных генов приводят к образованию мулатов с разной интенсивностью окраски кожи: от темной до светлой.
Различают две основных группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельнимы генами. Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие первичных и вторичных продуктов, которые обусловят тот или иной признак. В цитоплазме происходит взаимодействие между белками - ферментами, синтез которых опрелятся генами, или между веществами, которые образовываются под влиянием этих ферментов.
Возможны следующие типы взаимодействия:
1) для образования определенного признака необходимо взаимодействие двух ферментов, синтез которых опрелятся двумя неаллельнимы генами;
2) фермент, что был синтезирован с участием одного гена, полностью подавляет или инактивирует действие фермента, что был образован другим неаллельным геном;
3) два ферменты, образование которых контролируется двумя неаллельми генами, влияющими на один признак или на один процесс так, что их совместное действие приводит к возникновению и усилению проявления признака.
Взаимодействие аллельных генов
Гены, которые занимают идентичные (гомологические) локусы в гомологичных хромосомах, называются аллельными. У каждого организма есть по два аллельных гена.
Известны такие формы взаимодействия между аллельными генами: полное доминирование, неполное доминирование, кодоминированием и сверхдоминирование.
Основная форма взаимодействия - полное доминирование, которое впервые описано Г. Менделем. Суть его заключается в том, что в гетерозиготном организме проявление одной из аллелей доминирует над проявлением другой. При полном доминировании расщепления по генотипу 1:2:1 не совпадает с расщеплением по фенотипу - 3:1. В медицинской практике с двух тысяч моногенных наследственных болезней почти в половины имеет место доминированое проявления патологических генов над нормальными. В гетерозигот патологический аллель проявляется в большинстве случаев признаками заболевания (доминантный фенотип).
Неполное доминирование - форма взаимодействия, при которой у гетерозиготного организма (Аа) доминантный ген (А) не полностью подавляет рецессивный ген (а), вследствие чего проявляется промежуточный между родительскими признак. Здесь расщепление по генотипу и фенотипу совпадает и составляет 1:2:1
При кодоминировании в гетерозиготных организмах каждый из аллельных генов вызывает формирование зависимого от него продукта, то есть оказываются продукты обеих аллелей. Классическим примером такого проявления является система групп крови, в частности система АBО, когда эритроциты человека несут на поверхности антигены, контролируемые обеими аллелями. Такая форма проявления носит название кодоминированием.
Сверхдоминирование - когда доминантный ген в гетерозиготном состоянии проявляется сильнее, чем в гомозиготном. Так, у дрозофилы при генотипе АА-нормальная продолжительность жизни; Аа - удлиненная триватисть жизни; аа - летальный исход.
Множественный алелизм
У каждого организма есть только по два аллельных гена. Вместе с тем нередко в природе количество аллелей может быть более двух, если какой то локус может находится в разных состояниях. В таких случаях говорят о множественные аллели или множественный аллеломорфизм.
Множественные аллели обозначаются одной буквой с разными индексами, например: А, А1, А3 ... Аллельные гена локализуются в одинаковых участках гомологичных хромосом. Поскольку в кариотипе всегда присутствуют по две гомологичных хромосомы, то и при множественных аллелях каждый организм может иметь одновременно лишь по два одинаковых или различных аллели. В половую клетку (вместе с различием гомологичних хромосом) попадает только по одному из них. Для множественных аллелей характерное влияние всех аллелей на один и тот же признак. Отличие между ними заключается лишь в степени развития признака.
Второй особенностью является то, что в соматических клетках или в клетках диплоидных организмов содержится максимум по две аллели из нескольких, поскольку они расположены в одном и том же локусе хромосомы.
Еще одна особенность присуща множественным аллелям. По характеру доминирования аллеломорфные признаки размещаются в последовательном ряду: чаще нормальный, неизмененный признак доминирует над другими, второй ген ряда рецессивный относительно первого, однако доминирует над следующими и т.д. Одним из примеров проявления множественных аллелей у человека есть группы крови системы АВО.
Множественный алелизм имеет важное биологическое и практическое значение, поскольку усиливает комбинативну изменчивость, особенно генотипического.
Взаимодействие неалельних генов
Известно много случаев, когда признак или свойства детерминируются двумя или более неалельнимы генами, которые взаимодействуют между собой. Хотя и здесь взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими продукты. При этом имеет место отклонение от менделивских закономерностей расщепления.
Различают четыре основных типа взаимодействия генов: комплементарность, эпистаз, полимерию и модифицирующее действие (плейотропия).
Комплементарность это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причем соответственный признак развивается только в присутствии обоих неаллельних генов. Например, сера окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) - альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению гибридов в соотношении 9:3:4. Числовые соотношения при комплементарном взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка - интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз -это такое взаимодействие неаллельных генов, при котором один ген подавляет действие другого неаллельного гена. Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого розличают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При епистатичном взаимодействияи генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.