Zum ersten Mal taucht das Atmungssystem in marinen Anneliden auf - Sandwürmern und Nereiden, bei denen sich primitive Kiemen an den Rückenästen der Parapodien befinden. Darüber hinaus findet bei Anneliden der Gasaustausch durch die blutgefäßreiche Haut statt.
Bei Säugetieren und Weichtieren hängt die Struktur der Atmungsorgane von den Bedingungen ihres Lebensraums ab: Bei aquatischen Formen sind dies Kiemen, die im Wasser gelösten Sauerstoff verwenden können; in terrestrischen Lungen und Luftröhren, angepasst an die Verwendung von Luftsauerstoff.
Krebstiere haben also Kiemen, die sich an den Beinen und Kiefern unter den seitlichen Falten des Cephalothoracic-Schildes befinden, wo sie ständig von Wasser gewaschen werden.
Das Atmungssystem der Spinnentiere wird entweder durch blattförmige Lungen oder Luftröhren dargestellt. Beide öffnen sich nach außen mit speziellen Öffnungen - Narben an den Seitenteilen der Segmente. In den Lungensäcken befinden sich zahlreiche Blattfalten, in denen Blutkapillaren verlaufen. Die Lungen von Spinnentieren sind homolog zu den Kiemen von Krebstieren. Die Luftröhre ist ein System verzweigter Röhren, die direkt zu allen Organen führen, wo Gewebeaustausch stattfindet.
Insekten atmen durch die Luftröhre.
Bei vielen Arthropoden, die eine dünne Chitinhülle und eine relativ große Körperoberfläche haben, wird auch eine diffuse Atmung beobachtet.
Bei Weichtieren sind die Hauptatmungsorgane Kiemen, mit Ausnahme von Landmuscheln, beispielsweise einigen Schnecken, die Kiemen verloren haben und deren Mantelhöhle sich in eine Lunge verwandelt hat.
Viele Wirbellose verfügen über Vorrichtungen, die die Atmungsoberfläche in Form von lokal spezialisierten Atmungsorganen vergrößern.
Evolution des Atmungssystems in Akkordaten .
Das Atmungssystem aller Akkordaten ist sowohl dem Ursprung als auch der Topographie nach mit dem Entoderm verbunden.
Bei aquatischen Chordaten wird die Atmungsfunktion von Kiemenschlitzen übernommen, die den vorderen Abschnitt des Darmröhren-Pharynx durchdringen. Bei terrestrischen Chordaten schließen sich die Kiemenschlitze während der Embryonalentwicklung und verschwinden dann. Die Funktion der Atmung übernimmt die Lunge, die aus der Ausstülpung des Darmschlauches gebildet wird. Die Entwicklung des Kiemenapparates bei Chordatieren äußerte sich in einer Abnahme der Zahl der Kiemenschlitze bei gleichzeitiger Vergrößerung der Atemfläche durch die Bildung von Kiemenfäden.
Die Entwicklung der Lunge ging in Richtung der Isolierung der Atemwege und einer Vergrößerung der Atemoberfläche durch die Bildung von Lungen mit einer schwammigen Struktur mit einem komplexen System von Verzweigungen intrazellulärer Bronchien, die in Vesikeln mit Zellzellen enden.
Das primitivste Atmungssystem befindet sich in der Lanzette, die zu den unteren Akkordaten gehört (Subtyp Cranial). Der vordere Darm (Rachenwand) ist von Kiemenschlitzen (bis zu 150 Paar) durchzogen, die in die arterielle (periobranchiale) Höhle münden.
Bei Cyclostomen (Subtyp Wirbeltiere) sind die Atmungsorgane ebenfalls Kiemenschlitze, aber es gibt bereits weniger davon (5-15 Paare). Sie kommunizieren mit dem vorderen Darm und öffnen sich mit unabhängigen Öffnungen nach außen.
Echte Kiemen treten bei Fischen zwischen Akkordaten auf. Sie sind dünne Schleimhautfalten des Rachens, die auf den Kiemenbögen aufliegen und durch die Kiemenarterien, die hier in Kapillaren aufbrechen, mit venösem Blut versorgt werden. Fische haben 4-7 (normalerweise 5) Kiemensäcke zwischen den Kiemenbögen. Staubblätter auf der konvexen Oberfläche der Kiemenbögen verhindern, dass Nahrung aus dem Rachen in die Kiemen gelangt.
Fische haben neben Kiemen zusätzliche Atmungsorgane, die es ihnen ermöglichen, Luftsauerstoff zu nutzen. Ein solches Organ bei Fischen ist die Schwimmblase. Seine Wände sind reich an Blutgefäßen, daher kann es bei einigen Fischen, die sich in den Schlick graben, zum Gasaustausch dienen. Die Schwimmblase entwickelt sich bei den meisten Fischen aus den dorsalen Teilen des Pharynx und ist kein Homolog der Lunge. Nur bei Lappenflossenfischen ist die Schwimmblase als Ausstülpung des ventralen Teils des Pharynx ausgebildet und dient als Homolog der Landtierlunge, da sich die Lunge der Wirbeltiere aus dem ventralen Teil des Kiemensacks entwickelt.
Lungenfische haben Lungen, aber keine Schwimmblase.
Bei Amphibienlarven werden die Atmungsorgane wie bei Fischen durch baumverzweigte äußere Kiemen dargestellt. Bei den meisten erwachsenen Amphibien erscheinen Lungen in Form von dünnwandigen paarigen Auswüchsen des ventralen Pharynx hinter dem letzten Kiemensack. Aufgrund des Fehlens von Brust und Zwerchfell gelangt Luft aus der Mundhöhle aufgrund von Schluckbewegungen, die vom Kinn-Zungenbein-Muskel ausgeführt werden. Das Material der dem Hypoglossusbogen folgenden Kiemenbögen ist teilweise Teil des Kehlkopfknorpels, der erstmals bei Amphibien auftritt und das erste Organ ist, das mit den unteren Atemwegen in Verbindung steht. Die Lunge beginnt direkt am Kehlkopf. Sie sind grobmaschig und haben eine kleine Atmungsoberfläche und damit einen Gasaustausch mehr Sie erfolgt über die Haut, die von einer Vielzahl von Blutkapillaren durchzogen und mit Schleimdrüsen versorgt ist.
Bei Reptilien ist das Atmungssystem kompliziert. Ihre Lungen sind bereits feinzellig, sie enthalten zahlreiche Zellkreuze und haben eine große Atemfläche. In den Atemwegen werden im Zusammenhang mit dem endgültigen Austritt von Reptilien an Land Fortschritte beobachtet: Die oberen Atemwege werden unterschieden, obwohl sie nicht vollständig von der Mundhöhle abgegrenzt sind - dies ist die Nasenhöhle und die unteren sind der Kehlkopf. Luftröhre und Bronchien. Zum ersten Mal erscheint das Zwerchfell, das passiv an der Atmung teilnimmt. Es trennt entweder Brust- und Bauchhöhle teilweise oder ist frei von Muskelfasern.
Der Atemmechanismus wird durch Kontraktion der Zwischenrippenmuskeln ausgeführt, die den Brustkorb in Bewegung setzen.
Bei Vögeln sind die Lungen dichte schwammige Körper, die von Verzweigungen der Bronchien durchbohrt sind, und keine Beutel wie bei Reptilien. Außerdem werden sie als Luftspeicher mit dünnwandigen Airbags ergänzt. Letztere befinden sich zwischen allen Luftorganen, zwischen den Muskeln, in der Knochenhöhle und unter dem Knochen.
Wenn die Flügel angehoben werden, werden die Luftsäcke durch die Lungen mit Luft gefüllt, wenn die Flügel gesenkt werden, wird die Luft durch die Lungen freigesetzt. Vögel führen also während des Fluges eine Doppelatmung durch. In Ruhe atmet der Vogel nur durch Ausdehnen und Zusammenziehen der Brust.
Die Atmungsorgane aller Säugetiere sind durch die Komplexität sowohl der Lungen, die eine alveoläre Struktur haben, als auch der Atemwege gekennzeichnet. Es gibt eine weitere Verzweigung des Bronchialbaums im Vergleich zu den vorherigen Klassen von Wirbeltieren. Die Luftröhre ist in Bronchien unterteilt, die sich in Bronchien zweiter, dritter und vierter Ordnung verzweigen, und in die kleinsten selbst - Bronchiolen, an deren Zweigen sich Alveolen befinden, Lungenbläschen, die eine riesige Fläche im Beutel haben (etwa 90 cm im Quadrat), in dem Gasaustausch stattfindet. Der Hauptmuskel, der spielt essentielle Rolle beim Atmen ist das Zwerchfell. Die Atemwege von Säugetieren sind mit Flimmerepithel ausgekleidet und vollständig vom Verdauungssystem getrennt.
Es gibt zwei Arten von Atmungsorganen von Wirbeltieren – Kiemen und Lungen, und bei einem erheblichen Teil der Wirbeltiere ist die Haut für die Atmung unerlässlich. Der Kiemenapparat ist ein System paariger, normalerweise symmetrisch angeordneter Schlitze, die dazu dienen, den Pharynx mit der äußeren Umgebung zu verbinden. Die Vorder- und Hinterwände der Kiemenschlitze sind mit einer Schleimhaut ausgekleidet, die lamellare Auswüchse bildet; Auswüchse sind in Blütenblätter unterteilt, die Kiemen genannt werden. Jede Kiemenplatte über den Blütenblättern wird als Halbkieme bezeichnet. Zwischen den Kiemenschlitzen (in den Kiemensepten) befinden sich die viszeralen Kiemenbögen. Somit ist jeder Kiemenbogen mit zwei Halbkiemen zweier unterschiedlicher Kiemenschlitze verbunden. Die Atmungsorgane von Landwirbeltieren - die Lungen - in der Abbildung sind ein Paar Säcke, die sich durch die Kehlkopfspalte in den Pharynx öffnen. Embryonal entstehen die Lungen in Form einer Ausstülpung der Bauchwand des Pharynx im hinteren Kiemenapparat, sind also endodermalen Ursprungs. In den frühen Stadien der Embryonalentwicklung ähneln die Lungenknospen einem Paar innerer (endodermaler) Kiemenschlitze. Diese Umstände sowie die den Lungen und Kiemen gemeinsamen Blutversorgungs- und Innervationsmerkmale zwingen uns, die Lunge als Homologe des hinteren Kiemensackpaares zu betrachten. Die Haut ist an der Atmung beteiligt, wenn keine dichten Horn- oder Knochenschuppen darin vorhanden sind, beispielsweise bei Amphibien, Fischen mit nackter Haut. Funktionell ist das Atmungssystem an der Anreicherung des Blutes mit Sauerstoff und dem Abtransport von Kohlendioxid beteiligt. Ammoniak wird bei niederen Wassertieren über das Atmungssystem freigesetzt. Bei Warmblütern ist es an Prozessen der Thermoregulation beteiligt. Das Funktionsprinzip des Atmungssystems ist der Austausch von CO2 und O2 zwischen den gegeneinander gerichteten Gas- und Blutströmen. Die Atmung ist ein Prozess, der Sauerstoff aufnimmt und Kohlendioxid freisetzt. Das Atmungssystem erfüllt die wichtigste Funktion - den Gasaustausch, ohne den das Leben unmöglich ist, da die Energieumwandlung im Körper durch oxidativen Zerfall erfolgt Nährstoffe unter Beteiligung von Sauerstoff. Gasaustausch in der Lunge. Das Blut, das vom Herzen (venös) zur Lunge fließt, enthält wenig Sauerstoff und viel Kohlendioxid; die Luft in den Lungenbläschen hingegen enthält viel Sauerstoff und weniger Kohlendioxid. Als Ergebnis tritt eine Zwei-Wege-Diffusion durch die Wände der Alveolen und Kapillaren auf. Sauerstoff gelangt ins Blut und Kohlendioxid wandert aus dem Blut in die Alveolen. Im Blut gelangt Sauerstoff in die roten Blutkörperchen und verbindet sich mit Hämoglobin. Mit Sauerstoff angereichertes Blut wird arteriell und tritt durch die Lungenvenen in den linken Vorhof ein. Gasaustausch im Gewebe. Der Gasaustausch im Gewebe erfolgt in Kapillaren. Durch ihre dünnen Wände gelangt Sauerstoff aus dem Blut in die Gewebeflüssigkeit und dann in die Zellen, und Kohlendioxid aus den Geweben gelangt in das Blut. Die Sauerstoffkonzentration im Blut ist höher als in den Zellen, sodass er leicht in sie diffundiert. Die Konzentration von Kohlendioxid in den Geweben, in denen es gesammelt wird, ist höher als im Blut. Daher gelangt es ins Blut, wo es bindet Chemische Komponenten Plasma und teilweise mit Hämoglobin, wird über das Blut zur Lunge transportiert und in die Atmosphäre freigesetzt.
5.1. Entstehung und Funktion des Atmungssystems.
Das Atmungssystem tierischer Organismen ist endodermalen Ursprungs, da es ursprünglich damit assoziiert ist Verdauungssystem. Die Funktion der Atmungsorgane ist der Gasaustausch zwischen einem bestimmten Organismus und der Umgebung.
5.2. Die Entwicklung des Atmungssystems bei einer Reihe von Wirbellosen.
Zum ersten Mal taucht das Atmungssystem in marinen Anneliden auf - Sandwürmern und Nereiden, bei denen sich primitive Kiemen an den Rückenästen der Parapodien befinden. Darüber hinaus findet bei Anneliden der Gasaustausch durch die blutgefäßreiche Haut statt.
Bei Säugetieren und Weichtieren hängt die Struktur der Atmungsorgane von den Bedingungen ihres Lebensraums ab: Bei aquatischen Formen sind dies Kiemen, die im Wasser gelösten Sauerstoff verwenden können; in terrestrischen Lungen und Luftröhren, angepasst an die Verwendung von Luftsauerstoff.
Krebstiere haben also Kiemen, die sich an den Beinen und Kiefern unter den seitlichen Falten des Cephalothoracic-Schildes befinden, wo sie ständig von Wasser gewaschen werden.
Das Atmungssystem der Spinnentiere wird entweder durch blattförmige Lungen oder Luftröhren dargestellt. Beide öffnen sich nach außen mit speziellen Öffnungen - Narben an den Seitenteilen der Segmente. In den Lungensäcken befinden sich zahlreiche Blattfalten, in denen Blutkapillaren verlaufen. Die Lungen von Spinnentieren sind homolog zu den Kiemen von Krebstieren. Die Luftröhre ist ein System verzweigter Röhren, die direkt zu allen Organen führen, wo Gewebeaustausch stattfindet.
Insekten atmen durch die Luftröhre.
Bei vielen Arthropoden, die eine dünne Chitinhülle und eine relativ große Körperoberfläche haben, wird auch eine diffuse Atmung beobachtet.
Bei Weichtieren sind die Hauptatmungsorgane Kiemen, mit Ausnahme von Landmuscheln, beispielsweise einigen Schnecken, die Kiemen verloren haben und deren Mantelhöhle sich in eine Lunge verwandelt hat.
Viele Wirbellose verfügen über Vorrichtungen, die die Atmungsoberfläche in Form von lokal spezialisierten Atmungsorganen vergrößern.
5.3. Die Entwicklung des Atmungssystems in Akkordaten.
Das Atmungssystem aller Akkordaten ist sowohl ursprünglich als auch topographisch mit dem Darm, dh mit dem Endoderm, verbunden.
Bei aquatischen Chordaten wird die Atmungsfunktion von Kiemenschlitzen übernommen, die den vorderen Abschnitt des Darmröhren-Pharynx durchdringen. Bei terrestrischen Chordaten schließen sich die Kiemenschlitze während der Embryonalentwicklung und verschwinden dann. Die Funktion der Atmung übernimmt die Lunge, die aus der Ausstülpung des Darmschlauches gebildet wird.
Die Entwicklung des Kiemenapparates bei Chordatieren äußerte sich in einer Abnahme der Zahl der Kiemenschlitze bei gleichzeitiger Vergrößerung der Atemfläche durch die Bildung von Kiemenfäden.
Die Entwicklung der Lunge ging in Richtung der Isolierung der Atemwege und einer Vergrößerung der Atemoberfläche durch die Bildung von Lungen mit einer schwammigen Struktur mit einem komplexen System von Verzweigungen intrazellulärer Bronchien, die in Vesikeln mit Zellzellen enden.
Das primitivste Atmungssystem befindet sich in der Lanzette, die zu den unteren Akkordaten gehört (Subtyp Cranial). Der vordere Darmabschnitt (Pharynxwand) ist von Kiemenschlitzen (bis zu 150 Paar) perforiert, die in die arterielle (periobranchiale) Höhle münden.
Bei Cyclostomen (Subtyp Wirbeltiere) sind die Atmungsorgane ebenfalls Kiemenschlitze, aber es gibt bereits weniger davon (5-15 Paare). Sie kommunizieren mit dem vorderen Darm und öffnen sich mit unabhängigen Öffnungen nach außen.
Echte Kiemen treten bei Fischen zwischen Akkordaten auf. Sie sind dünne Schleimhautfalten des Rachens, die auf den Kiemenbögen liegen und durch die Kiemenarterien, die hier in Kapillaren aufbrechen, mit venösem Blut versorgt werden. Fische haben 4-7 (normalerweise 5) Kiemensäcke zwischen den Kiemenbögen. Staubblätter auf der konvexen Oberfläche der Kiemenbögen verhindern, dass Nahrung aus dem Rachen in die Kiemen gelangt.
Fische haben neben Kiemen zusätzliche Atmungsorgane, die es ihnen ermöglichen, Luftsauerstoff zu nutzen. Ein solches Organ bei Fischen ist die Schwimmblase. Seine Wände sind reich an Blutgefäßen, daher kann es bei einigen Fischen, die sich in den Schlick graben, zum Gasaustausch dienen. Die Schwimmblase entwickelt sich bei den meisten Fischen aus den dorsalen Teilen des Pharynx und ist kein Homolog der Lunge. Nur bei Lappenflossenfischen ist die Schwimmblase als Ausstülpung des ventralen Teils des Pharynx ausgebildet und dient als Homolog der Landtierlunge, da sich die Lunge der Wirbeltiere aus dem ventralen Teil des Kiemensacks entwickelt.
Lungenfische haben Lungen, aber keine Schwimmblase.
Bei Amphibienlarven werden die Atmungsorgane wie bei Fischen durch baumverzweigte äußere Kiemen dargestellt. Bei den meisten erwachsenen Amphibien erscheinen Lungen in Form von dünnwandigen paarigen Auswüchsen des ventralen Pharynx hinter dem letzten Kiemensack. Aufgrund des Fehlens von Brust und Zwerchfell gelangt Luft aus der Mundhöhle aufgrund von Schluckbewegungen, die vom Kinn-Zungenbein-Muskel ausgeführt werden. Das Material der dem Hypoglossusbogen folgenden Kiemenbögen ist teilweise Teil des Kehlkopfknorpels, der erstmals bei Amphibien auftritt und das erste Organ ist, das mit den unteren Atemwegen in Verbindung steht. Die Lunge beginnt direkt am Kehlkopf. Sie sind grobmaschig und haben eine kleine Atmungsfläche, daher erfolgt der Gasaustausch verstärkt über die Haut, die von einer Vielzahl von Blutkapillaren durchzogen und mit Schleimdrüsen versorgt ist.
Bei Reptilien ist das Atmungssystem kompliziert. Ihre Lungen sind bereits feinzellig, sie enthalten zahlreiche Zellkreuze und haben eine große Atemfläche. In den Atemwegen werden im Zusammenhang mit dem endgültigen Austritt von Reptilien an Land Fortschritte beobachtet: Die oberen Atemwege werden unterschieden, obwohl sie nicht vollständig von der Mundhöhle abgegrenzt sind - dies ist die Nasenhöhle und die unteren sind der Kehlkopf. Luftröhre und Bronchien. Zum ersten Mal erscheint das Zwerchfell, das passiv an der Atmung teilnimmt. Es trennt entweder Brust- und Bauchhöhle teilweise oder ist frei von Muskelfasern.
Der Atemmechanismus wird durch Kontraktion der Zwischenrippenmuskeln ausgeführt, die den Brustkorb in Bewegung setzen.
Bei Vögeln sind die Lungen dichte schwammige Körper, die von Verzweigungen der Bronchien durchbohrt sind, und keine Beutel wie bei Reptilien. Außerdem werden sie als Luftspeicher mit dünnwandigen Airbags ergänzt. Letztere befinden sich zwischen allen Luftorganen, zwischen den Muskeln, in der Knochenhöhle und unter dem Knochen.
Wenn die Flügel angehoben werden, werden die Luftsäcke durch die Lungen mit Luft gefüllt, wenn die Flügel gesenkt werden, wird die Luft durch die Lungen freigesetzt. Vögel führen also während des Fluges eine Doppelatmung durch. In Ruhe atmet der Vogel nur durch Ausdehnen und Zusammenziehen der Brust.
Die Atmungsorgane aller Säugetiere sind durch die Komplexität sowohl der Lungen, die eine alveoläre Struktur haben, als auch der Atemwege gekennzeichnet. Es gibt eine weitere Verzweigung des Bronchialbaums im Vergleich zu den vorherigen Klassen von Wirbeltieren. Die Luftröhre ist in Bronchien unterteilt, die sich in Bronchien zweiter, dritter und vierter Ordnung verzweigen, und in die kleinsten selbst - Bronchiolen, an deren Zweigen sich Alveolen befinden, Lungenbläschen, die eine riesige Fläche im Beutel haben (etwa 90 cm im Quadrat), in dem Gasaustausch stattfindet. Der Hauptmuskel, der beim Atmen eine wichtige Rolle spielt, ist das Zwerchfell. Die Atemwege von Säugetieren sind mit Flimmerepithel ausgekleidet und vollständig vom Verdauungssystem getrennt.
5.4. Anomalien und Fehlbildungen des Atmungssystems beim Menschen.
1. In der menschlichen Embryogenese spiegelt sich die anfängliche Verbindung zwischen Verdauungs- und Atmungssystem wider. In dieser Hinsicht wird deutlich, dass eine Person eine große Gruppe von angeborenen Fehlbildungen der Speiseröhre und Luftröhre des Typs hat ösophagotracheale Fisteln (« Kiemenspalten»).
2. Dysontogenetische bronchopulmonale Zysten- ein abgerundeter Streifen in der Lunge, der durch eine primitiv konstruierte Wand eines undifferenzierten Bronchus vom umgebenden Gewebe abgegrenzt ist. Diese Anomalien können durch eine Verletzung der alveolären Differenzierung des Lungengewebes erklärt werden.
3. Zystische Hypoplasie- Unterentwicklung der Lunge. Bei dieser Pathologie sind ganze Lungenlappen unterentwickelt, bei denen es sich um zahlreiche Hohlräume handelt, die mit großen Bronchien verbunden sind und eine kleine Oberfläche haben. Der Gasaustausch in solchen Bereichen der Lunge ist stark geschwächt.
4. Hypoplasie des Zwerchfells- Unterentwicklung des Zwerchfells von kleinen Defekten in seiner Kuppel bis zur vollständigen Aplasie. Diese Anomalie ist mit dem Leben nicht vereinbar und tritt häufiger zusammen mit anderen multiplen Fehlbildungen auf.
Vortrag 12
Klasse Amphibien (Amphibien)
Klassenmerkmal
Amphibien sind die ersten Wirbeltiere, die an Land landen, haben aber den Kontakt zu ihnen nicht verloren aquatische Umgebung. Die Fortpflanzung erfolgt im Wasser, es gibt eine aquatische fischähnliche Larve. Darüber hinaus ist für die meisten Amphibienarten auch im Erwachsenenalter eine enge Beziehung zum Wasser charakteristisch, sodass sie sowohl für das Leben im Wasser als auch an Land angepasst sind.
Abdeckungen dargestellt durch Haut, weich, nackt, durchlässig für Gase und Wasser.
Skelett und Muskeln . Das Skelett erfährt eine Reihe von Transformationen im Zusammenhang mit Boden Weg Leben. Die zervikalen und sakralen Abschnitte der Wirbelsäule sind getrennt und haben jeweils einen Wirbel. Der Schädel ist mit Hilfe von zwei Kondylen beweglich am Halswirbel befestigt. Aus den paarigen Flossen von Süßwasserfischen mit Lappenflossen für die Bewegung an Land werden Gliedmaßen gebildet, die ein System von Hebeln darstellen. Die Muskulatur verliert ihre metamere Struktur und wird durch viele einzelne Muskeln repräsentiert.
Atmungssystem . Das Atmungssystem von Amphibien ist insofern interessant, als im Entwicklungsprozess ein Übergang von der Kiemenatmung zur Lungenatmung stattfindet. Die Hautatmung ist von großer Bedeutung. Die Atemwege sind schwach entwickelt.
Kreislauf . Seit dem Erscheinen der Lungen tritt ein (kleiner) Lungenkreislauf auf. Das Amphibienherz wird dreikammerig, drei Paare von Arterienbögen gehen davon aus. Der Stoffwechsel ist noch nicht sehr intensiv, Amphibien sind nicht in der Lage, eine konstante Körpertemperatur aufrechtzuerhalten, sie gehören zu den wechselwarmen Tieren.
Nervensystem . Im Gehirn sind die großen Hemisphären des Vorderhirns vergrößert und vollständig getrennt. Mittelhirn und Kleinhirn sind leicht entwickelt. 10 Hirnnervenpaare verlassen das Gehirn.
Das Leben in der Luft hat zum Auftreten einer Reihe von Merkmalen in den Sinnen geführt. Aufgrund der konvexen Hornhaut und der abgeflachten Linse verbessert sich die Akkommodation. Um die Augen vor Verstopfung und Austrocknung zu schützen, haben Amphibien bewegliche Augenlider und eine Nickhaut. Um Geräusche in der Luft wahrzunehmen, erscheint das Trommelfell, dahinter befindet sich die Lufthöhle des Mittelohrs und ein Gehörknochen - der Steigbügel, der Schwingungen zum Innenohr leitet. Die Eustachische Röhre verbindet die Mittelohrhöhle mit der Mundhöhle. Choanae erscheinen, innere Nasenlöcher, Nasengänge werden durch.
Verteiler . In der modernen Fauna gibt es mehr als 4.000 Amphibienarten, die in die Ordnungen Schwanzlose (Anura), Schwanzlose (Caudata) und Beinlose (Apoda) unterteilt sind.
Die folgenden Aromorphosen trugen zur Entstehung von Amphibien an Land bei: 1. Licht und Lungenatmung traten auf. 2. Machte es schwieriger Kreislauf, der Lungenkreislauf hat sich entwickelt, das heißt, Amphibien haben zwei Blutkreislaufkreise - einen großen und einen kleinen. Das Herz wurde dreikammerig. 4. Es wurden paarige, fünffingrige Gliedmaßen gebildet, die ein System von Hebeln mit Gelenkverbindungen sind und für die Bewegung an Land ausgelegt sind. 5. In der Wirbelsäule erschien eine Halsregion, die die Bewegung des Kopfes sicherstellte, und eine Sakralregion, der Ort der Befestigung des Beckengürtels. 6. Erschien das Mittelohr, Augenlider, Choanen.
Struktur und Leben . Schwanzamphibien haben einen länglichen Körper, einen langen Schwanz und Gliedmaßen. Schwanzlose haben einen kurzen Körper, der in dorsal-abdominaler Richtung abgeflacht ist, der Schwanz fehlt. Die Beinlosen haben einen länglichen, wurmartigen Körper mit zahlreichen ringförmigen Falten nackter Haut, keine Gliedmaßen und einen kurzen Schwanz.
Das Integument wird durch eine mehrschichtige Epidermis und Dermis dargestellt. Die Haut ist weich, nackt, gas- und wasserdurchlässig und von einem Netz von Blutgefäßen durchzogen. Es fungiert bei Amphibien als zusätzliches Atmungsorgan und enthält daher zahlreiche Drüsen, die ständig Schleim absondern. Die Durchfeuchtung der Haut beruht darauf, dass nur durch den Wasserfilm ein Gasaustausch zwischen Körper und Umgebung stattfinden kann. Neben Schleimhäuten gibt es auch Giftdrüsen.
Phylogenie . Devon Paläozoikum, in dem die ersten Amphibien auftauchten, war anscheinend von saisonalen Dürren geprägt, in denen das Leben in vielen Süßwasserkörpern für Fische schwierig war. Die Erschöpfung des Wassers mit Sauerstoff und die Schwierigkeit, darin zu schwimmen, wurde durch die verfallenden Überreste einer üppigen Vegetation erleichtert, die in der Karbonzeit in Sümpfen und an den Ufern von Stauseen wuchs. Unter diesen Bedingungen könnten einige Fische an die Oberfläche steigen und Luft schlucken, es bildeten sich Lungensäcke. Aber mit einer starken Austrocknung der Stauseen wurde das Leben für Fische bereits unmöglich. Sie konnten sich nicht an Land bewegen und starben. Nur die aquatischen Wirbeltiere, die neben der Fähigkeit zur Lungenatmung auch an Land bewegliche Gliedmaßen erwarben, konnten diese Bedingungen überleben. Sie krochen an Land und überquerten benachbarte Stauseen, in denen noch Wasser aufbewahrt wurde.
Der Übergang der Wirbeltiere von einer aquatischen zu einer terrestrischen Lebensweise und das Auftreten von Amphibien wurde von zwei entscheidenden Anpassungen begleitet: Atmen von Luftsauerstoff und Fortbewegen auf festem Untergrund. Mit anderen Worten, die Kiemenatmung hätte durch die Lungenatmung ersetzt werden sollen und die Flossen durch fünffingrige Gliedmaßen, die ein System von Hebeln sind, die dazu dienen, den Körper zu stützen und sich zu bewegen. Parallel dazu veränderten sich auch andere Organsysteme. Die meisten Forscher glauben, dass Amphibien von Süßwasser-Buschflossen abstammen. ripidistisch Fisch vor 370-350 Millionen Jahren. Erstmals erschienen ichthyostegidae, Dann Stegocephalianer- muschelköpfig. Die ersten Amphibien, die am Ende des Devons im Süßwasser auftauchten, waren Ichthyostegiden. Sie waren echte Übergangsformen zwischen Lappenflossenfischen und Amphibien. Sie hatten also die Rudimente des Kiemendeckels, einen echten Fischschwanz, die Haut war mit kleinen Fischschuppen bedeckt. Gleichzeitig hatten sie jedoch fünffingrige Gliedmaßen von Landwirbeltieren gepaart. Ichthyostegiden lebten nicht nur im Wasser, sondern auch an Land. Im Allgemeinen starben alle paläozoischen Amphibien während der Trias aus.
Merkmale der Struktur und Reproduktion.
Machen wir uns am Beispiel eines Frosches mit den Merkmalen der Struktur vertraut. Der Körper ist in dorsal-abdominaler Richtung abgeflacht, besteht aus Kopf, Rumpf und Gliedmaßen. Die nackte Haut ist feucht, hyperosmotisch, Wasser und ein Teil des Sauerstoffs gelangt durch die Haut in den Körper des Frosches. Auf dem Kopf sind hervorstehende Augen, in Verbindung mit dem Leben in der Luft, Augenlider und eine Nickhaut erschienen. Am Ende der Schnauze befinden sich die Nasenlöcher, die Nasengänge öffnen sich mit Choans in die Mund-Rachen-Höhle. Hinter den Ohröffnungen bei Männchen vieler Arten befinden sich aufblasende Blasen - Resonatoren, mit deren Hilfe die abgegebenen Geräusche verstärkt werden. Die Halsregion ist schwach ausgeprägt, der Schwanz fehlt. Hebelartige Gliedmaßen, hintere Fünffingergliedmaßen sind länger, vordere Vierfingerglieder. Die Farbgebung ist herablassend.
Skelett und Muskeln . Das Skelett eines Frosches ist wie das aller Wirbeltiere in vier Abschnitte unterteilt: das Achsenskelett, das Schädelskelett, das Gliedmaßenskelett und das Skelett der Gliedmaßengürtel.
Das Achsenskelett wird durch die Wirbelsäule dargestellt, in der neben den für Fische charakteristischen Rumpf- und Schwanzabschnitten auch die Hals- und Kreuzbeinabschnitte auftraten. Der Schädel eines Frosches ist mit Hilfe von zwei Kondylen mit einem einzigen Halswirbel beweglich gegliedert, was die Bewegung des Kopfes in einer vertikalen Ebene gewährleistet (der Kopf kann sich nicht in einer horizontalen Ebene bewegen).
Die Anzahl der Wirbel der Rumpfregion kann unterschiedlich sein, die größte Anzahl ist bei beinlosen Amphibien (mehr als hundert). Anuran-Amphibien haben die wenigsten Wirbel im Rumpfbereich, der Frosch hat sieben davon. Der Frosch hat keine Rippen, aber bei Amphibien mit Schwanz entwickeln sich kurze obere Rippen an den Wirbeln des Rumpfes, und beinlose haben echte Rippen. Die Sakralregion umfasst einen Wirbel, der lange Querfortsätze trägt, an denen die Beckenknochen des Beckens befestigt sind. Der Schwanzabschnitt des Frosches endet mit einem Steißbein - Urostyle - einem Knochen, bei dem es sich um mehrere Wirbel handelt, die im Verlauf der Embryonalentwicklung verschmolzen sind. Im Zusammenhang mit der Landung kam es zu Veränderungen des Schädels sowie zu einer Verringerung der Kiemen und des Kiemenapparates.
Reis. 146. Froschskelett:
1 - Halswirbel; 2 - Sakralwirbel; 3 - Urostil; 4 - Brustbein; 5 - Krähenknochen (Coracoid); 6 - Schlüsselbein; 7 - Schulterblatt; 8 - Sitzbein; 9 - Darmbein; 10 - Schamknorpel; 11 - Oberarmknochen; 12 - Unterarm (Radius und Elle); 13 - Bürste; 14 - Oberschenkelknochen; 15 - Unterschenkelknochen (Schienbein und Schienbein); 16 - Fußwurzel; 17 - Mittelfuß; 18 - Phalangen der Finger.
Da das spezifische Körpergewicht an Land deutlich größer ist als im Wasser, haben Amphibien Gliedmaßen, die in beweglich verbundene Abschnitte unterteilt sind. Eine solche Struktur reduziert den Aufwand, der aufgewendet wird, um den Körper unter irdischen Bedingungen zu halten und zu bewegen, erheblich. Die Vorderbeine, normalerweise mit vier Fingern (der erste Finger ist reduziert), bestehen aus drei Abschnitten: der Schulter - dem Humerus, dem Unterarm - dem verschmolzenen Radius und der Ulna und der Hand, dargestellt durch die Knochen des Handgelenks, der Mittelhand und der Phalangen der Finger.
Die Hinterbeine bestehen aus drei Abschnitten: Oberschenkel, Unterschenkel und Fuß. Der Oberschenkel besteht aus dem Femur, der Unterschenkel aus der verschmolzenen Tibia und Tibia, der Fuß aus den Knochen von Tarsus, Metatarsus und Phalangen der Finger.
Der Schultergürtel (Abb. 145) des Frosches umschließt den Körper mit einem weiten Halbkreis und ist in den Muskeln fixiert. Es wird durch mehrere gepaarte Knochen dargestellt: Schulterblätter, die in sh enden

Reis. 145. Froschschultergürtel:
1 - Prästernum; 2 - Schlüsselbein; 3 - Coracoid (Krähenknochen); 4 - Brustbein; 5 - Schulterblatt und supraskapularer Knorpel.
mit großen supraskapularen Knorpeln, Krähenknochen und Schlüsselbeinen sowie einem ungepaarten Knochen - dem Brustbein (Abb. 146). Der Beckengürtel besteht aus drei gepaarten Knochen, die aufgrund schwerer Belastungen verschmolzen sind: Becken-, Scham- und Sitzbeinknochen. Mit Hilfe des Darmbeins wird der Beckengürtel an den Querfortsätzen der Kreuzbeinwirbel befestigt.
Aufgrund komplexerer Bewegungen ist das Muskelsystem bei Amphibien viel komplexer als bei Fischen. Kräftige und komplex organisierte Muskeln der Vorder- und Hinterbeine entwickeln sich. Die für Fische charakteristische Muskelmetamerie (segmentierte Struktur) ist bei Amphibien gestört, der Aufbau der Muskulatur wird differenzierter.
Verdauungssystem
gekennzeichnet durch den Erwerb bestimmter Merkmale, die mit einer irdischen Lebensweise verbunden sind. Im Gegensatz zu Fischen entwickeln Amphibien Speicheldrüsen, deren Kanäle in die Mund-Rachen-Höhle münden. Außerdem erscheint eine Zunge, die über eigene Muskeln verfügt und an der Erfassung und Speicherung von Nahrung beteiligt ist. 
Reis. 147. Froschkopf:
1 - Kehlkopfspalte; 2 - Sprache; 3, 4 - innere bzw. äußere Nasenlöcher; 5 - Augen; 6 - Trommelfell.
Die Zunge befindet sich am Boden der Mund-Rachen-Höhle und kann eine andere Form haben. Bei einem Frosch kann der Zungenrücken nach vorne geworfen werden und dient als Werkzeug zum Fangen kleiner Tiere (Abb. 147). Die kleinen Zähne am Oberkiefer, die eine konische Form und leicht gebogene Rückenspitzen haben, helfen auch, Nahrung zu halten; Das Schieben des Nahrungsbolus wird durch die Augäpfel erleichtert, die nur durch eine dünne Schleimhaut vom Mund-Rachen-Raum getrennt sind und mit Hilfe spezieller Muskeln in den Mund-Rachen-Raum gezogen werden können.
Abb.148. Diagramm der Struktur der Froschlunge.
Durchsuchen vom Oropharynx durch eine kurze Speiseröhre gelangt in den Magen. Der Darm ist relativ länger als der von Fischen und besteht aus drei Abschnitten: vorderer, mittlerer und hinterer. Das Rektum mündet in die Kloake. Die große Leber sondert Galle ab, die sich in der Gallenblase ansammelt und durch die Gänge in den vorderen Teil des Dünndarms (den sogenannten Zwölffingerdarm) gelangt, wo auch die Pankreasgänge münden.
Atmungssystem . Amphibienlarven atmen mit Hilfe verzweigter äußerer Kiemen, die bei den meisten Arten nach der Metamorphose verschwinden. Bei erwachsenen Amphibien erfolgt der Gasaustausch über Haut und Lunge. Der Wert der Hautatmung ist sehr hoch, beispielsweise treten bei einem grünen Frosch 51% Sauerstoff durch die Haut ein und 86% Kohlendioxid werden freigesetzt.
Die Lungen der Amphibien sind Hohlsäcke mit mehr oder weniger ausgeprägter Zellstruktur (Abb. 148). Die Lungenoberfläche ist sehr klein und ihr Verhältnis zur Hautoberfläche beträgt 2:3. Auch die Atemwege sind schlecht entwickelt. Bei einem Frosch werden sie nur durch eine kurze Tracheal-Larynx-Kammer dargestellt, und bei Vertretern von Caudates werden sie durch eine lange Röhre - die Trachea - dargestellt.
Als Brustkorb fehlt, ist der Atmungsmechanismus von einem primitiven erzwungenen Typ. Die Inhalation erfolgt durch die Nasenlöcher, während der Mundboden abgesenkt wird, dann werden die Nasenlöcher mit Ventilen verschlossen, der Mundboden steigt wieder an und die Luft wird in die Lunge gedrückt. Die Ausatmung erfolgt mit Hilfe der Bauchmuskeln. Neben der Lunge findet ein zusätzlicher Gasaustausch in der Mundhöhle statt, deren Wand von einem Netz von Kapillaren durchzogen ist.
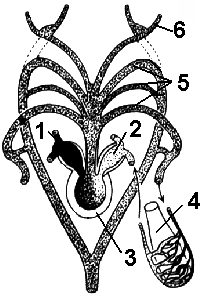
Reis. 149. Schema der Struktur des Herzens und der Arterienbögen:
1 - rechter Vorhof; 2 - linker Vorhof; 3 - Ventrikel; 4 - Lunge; 5 - Arterienbögen; 6 - Halsschlagadern.
ov.
Kreislauf unterteilt in zwei Kreisläufe des Blutkreislaufs: groß und klein (pulmonal). Das Herz ist dreikammerig und besteht aus zwei Vorhöfen und einem Ventrikel. Arterielles Blut tritt von der Lunge durch die Lungenvenen in den linken Vorhof ein, und gemischtes Blut tritt in den rechten Vorhof ein, da die Hohlvene aus innere Organe venöses Blut tritt ein, und die Hautvenen bringen arterielles Blut (Abb. 149).
Im Ventrikel vermischt sich das Blut aufgrund spezieller Teilungsmechanismen (verschiedene Auswüchse und eine Spiralklappe des Arterienkegels) nur teilweise.
Großer Kreislauf des Blutkreislaufs. Aus dem Ventrikel tritt Blut in den Arterienkegel ein, der sich weiter in drei Paare arterieller Gefäße verzweigt. Bei der Kontraktion der Herzkammer wird zunächst venöses Blut herausgedrückt, das die ersten beiden Arterienpaare füllt. Blut mit maximalem Sauerstoffgehalt tritt in das dritte Arterienpaar ein, von dem die Halsschlagadern abgehen und das Gehirn mit Blut versorgen.
Die Aortenbögen, die einen Halbkreis beschrieben haben, verschmelzen miteinander und bilden einen gemeinsamen Stamm der dorsalen Aorta, der die inneren Organe mit gemischtem Blut versorgt. Dann strömen venöses Blut (von den inneren Organen durch die Hohlvene) und arterielles (durch die Hautvenen) in den rechten Vorhof.
Die Lungenarterien transportieren sauerstoffarmes Blut in die Lunge, wo ein Gasaustausch stattfindet, dann gelangt arterielles Blut durch die Lungenvenen in den linken Vorhof - dies ist der Lungenkreislauf. Große Äste gehen von jeder Pulmonalarterie aus - Hautarterien, die Blut zur Haut transportieren, wo es oxidiert wird und dann in den rechten Vorhof eintritt.
Erythrozyten in Amphibien sind groß, bikonvex, haben einen Kern. Der Stoffwechsel ist höher als der von Fischen, aber nicht hoch genug, um eine konstante Körpertemperatur aufrechtzuerhalten, weshalb Amphibien zu den wechselwarmen Tieren gezählt werden.
H
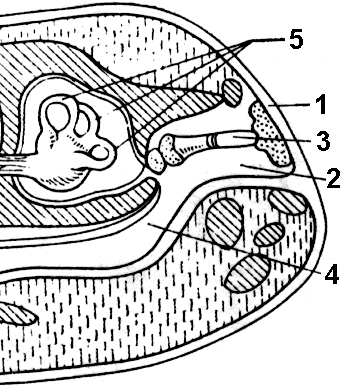
Abbildung 150. Querschnitt durch den Froschkopf im Ohrbereich:
1 - Trommelfell; 2 - Mittelohrhöhle; 3 - Steigbügel; 4 - Eustachische Röhre; 5 - Bogengänge.
Nervensystem und Sinnesorgane. Das Gehirn von Amphibien besteht wie das aller Wirbeltiere aus fünf Abschnitten: Vorderhirn, Zwischenhirn, Mittelhirn, Kleinhirn und Medulla oblongata. Im Vergleich zu Fischen hat bei Amphibien die relative Größe des Vorderhirns zugenommen und es hat eine vollständige Trennung seiner Hemisphären gegeben. Außerdem nahm die Zahl der Nervenzellen (graue Substanz) zu, die jedoch in den tiefen Schichten des Vorderhirns enthalten sind und an seiner Oberfläche fehlen. Das Mittelhirn ist relativ klein und das Kleinhirn aufgrund geringer Beweglichkeit und monotoner Bewegungen unterentwickelt. Die Medulla oblongata geht in das Rückenmark über.
Hörorgan. Neben dem Innenohr, das auch bei Fischen vorkommt, haben Amphibien ein entwickeltes Mittelohr, in dessen Höhle sich ein Steigbügel befindet - ein Gehörknochen, der erstmals bei Wirbeltieren auftauchte.
Um Schallschwingungen zu übertragen, liegt der Steigbügel an einem Ende am Trommelfell an, das die Mittelohrhöhle von der Außenumgebung trennt (Abb. 150), und am anderen Ende am ovalen Fenster, einem verdünnten Abschnitt des Septums zwischen Mittel- und Innenohr. Die Mittelohrhöhle ist durch einen engen Kanal mit der Mundhöhle verbunden - der Eustachischen Röhre. Dies ist notwendig, damit der Außen- und Innendruck auf das Trommelfell gleich sind, was die Membran vor einem Bruch schützt.
Das Sehorgan ist für Amphibien notwendig, um Objekte zu untersuchen, hauptsächlich an Land. Die dafür notwendigen Anpassungen äußern sich in der konvexen Form der Hornhaut und in der Linse, die als bikonvexe Linse ausgebildet ist (im Gegensatz zu Fischen, die eine sphärische Linse haben). Um die Augen vor Verstopfung und Austrocknung zu schützen, haben Amphibien außerdem bewegliche Augenlider, Nickhäute und Drüsen, die die Hornhaut mit Feuchtigkeit versorgen. Die Akkommodation erfolgt, wie bei Fischen, nur aufgrund der Bewegung der Linse.
Das Riechorgan der Amphibien befindet sich in gepaarten Riechkapseln, die durch die äußeren Nasenlöcher mit der äußeren Umgebung und mit der Mund-Rachen-Höhle kommunizieren - durch die Choanae dient das beschriebene System also nicht nur der Wahrnehmung von Gerüchen, sondern auch für Atmung. Die Seitenlinie ist charakteristisch für alle Amphibienlarven und befindet sich in der Haut, aber nicht in einer vertieften Rinne wie bei Fischen, sondern oberflächlich. In erwachsenen Formen bleibt die Seitenlinie nur bei Amphibien mit Wasserschwanz und einigen Anuren, auch Wassertieren, erhalten.
Ausscheidungssystem , wie bei Fischen, wird es durch zwei Rumpfnieren dargestellt, deren Funktion darin besteht, überschüssiges Wasser zu entfernen. Das Hauptausscheidungsprodukt ist Harnstoff. Die Harnleiter führen den Urin zur Kloake und dann zur Blase. Nach dem Füllen wird der Urin wieder in die Kloake und dann wieder ausgeschieden. Bei Männern erfüllt der Harnleiter eine andere Funktion - die Funktion des Vas deferens.
Amphibien sind gegenüber Süßwasser hyperosmotische Tiere. Dadurch gelangt ständig Wasser durch die Haut in den Körper, die wie bei anderen Landwirbeltieren keine Mechanismen besitzt, dies zu verhindern. Meerwasser ist in Bezug auf den osmotischen Druck im Gewebe von Amphibien hyperosmotisch, Wasser verlässt den Körper durch die Haut. Dies ist der wichtigste Grund, warum Amphibien nicht im Meerwasser leben können und darin an Austrocknung sterben.
Reproduktion und Entwicklung . Geschlechtsunterschiede zwischen Weibchen und Männchen von Amphibien werden meistens schlecht ausgedrückt. Bei den meisten Anuren sind die Männchen etwas kleiner als die Weibchen. Männliche Molche haben eine helle Paarungsfärbung. Es gibt auch Änderungen morphologischen Charakter Beispielsweise wächst beim männlichen Kammmolch während der Brutzeit ein ledriger Rand auf dem Rücken und dem Schwanz, und bei den Männchen schwanzloser Amphibien bildet sich an der Innenseite der Vorderbeine eine Brutschwiele.
Die Befruchtung kann sowohl innerlich als auch äußerlich erfolgen.
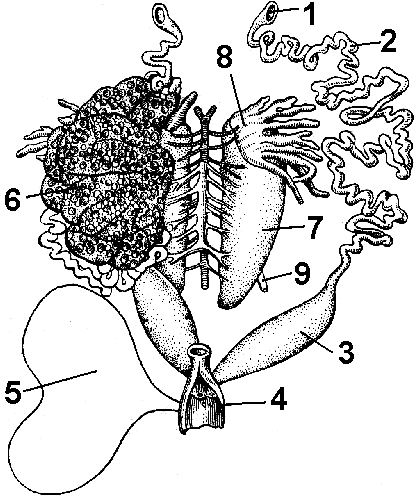
Reis. 151. Urogenitalsystem des weiblichen Frosches:
1 - Trichter des Eileiters; 2 - Eileiter (Müller-Kanal); 3 - Gebärmutter; 4 - Kloake; 5 - Blase; 6 - Eierstock; 7 - Niere; 8 - Fettkörper; 9 - Harnleiter.
hässlich. Die Entwicklung von Amphibien erfolgt mit Metamorphose, dh Larven entstehen aus Eiern, die sich normalerweise im Wasser entwickeln. Sie ernähren sich von Nahrung, die sich von der Ernährung Erwachsener unterscheidet, und unterscheiden sich von Erwachsenen in einigen fischähnlichen Strukturmerkmalen.
Die Geschlechtsorgane des männlichen Frosches werden durch gepaarte Hoden dargestellt. Die Hodenkanälchen münden in den Harnleiter (Wolfskanal). Die Befruchtung beim Frosch erfolgt äußerlich bzw. das Männchen hat keine Begattungsorgane.
Das Fortpflanzungssystem des Weibchens wird durch paarige Eierstöcke und Eileiter (Müller'sche Kanäle) dargestellt, die wie stark gewundene Röhren aussehen, die an einem Ende in die Kloake münden und am anderen Trichter tragen, in die die Eier fallen (Abb. 151).
Das Eigelb im Ei ist ungleichmäßig verteilt, es konzentriert sich auf das untere vegetative Plus einer helleren Farbe. Gegenüber (Tierpol) Eier enthalten
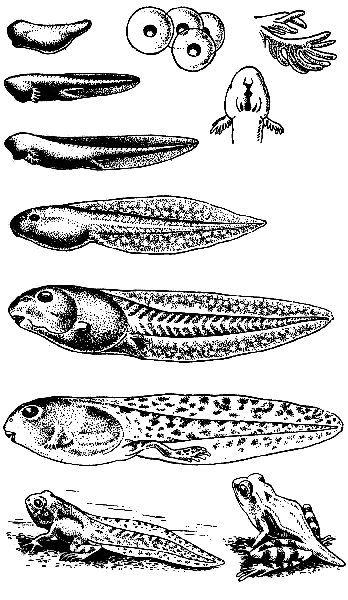
Abb.152. Froschentwicklung.
es ist ein Pigment, das ihm eine dunklere Farbe verleiht, was zur besseren Erwärmung dient.
Das Ei erfährt eine vollständige und ungleichmäßige Zerkleinerung. 8-10 Tage nach der Befruchtung durchbricht der Froschembryo die Eischalen und kommt in Form einer Larve – einer Kaulquappe – heraus.
Die Kaulquappe hat zunächst eine fischartige Struktur: Sie hat keine paarigen Gliedmaßen, das einzige Bewegungsorgan ist der Schwanz mit einer gut entwickelten Membran. Seine Atmungsorgane sind 2-3 Paar äußere Kiemen (Abb. 152), die Seitenlinie ist gut entwickelt, es gibt eine Sehne, vier Arterienbögen. In diesem Entwicklungsstadium hat die Kaulquappe ein zweikammeriges Herz und nur einen Blutkreislauf, wie bei Fischen.
Mit der weiteren Entwicklung verkümmern die äußeren Kiemen und es bilden sich Kiemenschlitze mit Blütenblättern, die dann mit der Lungenbildung auch verschwinden. Parallel dazu laufen auch andere Prozesse ab: Die Seitenlinie verschwindet, der Schwanz atrophiert, paarige Gliedmaßen entwickeln sich, die Notochord verschwindet und der Darm verkürzt sich. Die für Erwachsene charakteristische Kaulquappe geht von pflanzlicher Nahrung zu tierischer Nahrung über und verwandelt sich in einen Frosch.
Bestellen Sie Schwanzlurche (Caudata). Die älteste und relativ kleine Gruppe von Amphibien - etwa 340 Arten. Sie sind hauptsächlich in der gemäßigten Zone der westlichen und östlichen Hemisphäre verbreitet. Der Körper ist länglich, abgerundet und hat einen langen Schwanz, der das ganze Leben über bestehen bleibt. Die Vorder- und Hinterbeine sind gleich lang, sie bewegen sich also durch Kriechen oder Gehen. Unterarm und Unterschenkel haben einen typischen Aufbau und bestehen aus zwei Knochen, der Elle und Speiche, dem Schien- und Schienbein. Merkmale Interne Struktur sind die primitivsten für die Klasse als Ganzes. In den primitivsten Formen bleibt ein rudimentärer Akkord das ganze Leben lang erhalten. Es gibt rudimentäre obere Rippen.
Einige Vertreter, wie Proteus, behalten lebenslang äußere Kiemen. Die Rolle der Hautatmung ist groß. Die Mittelohrhöhle und das Trommelfell fehlen, bei einigen Arten bleibt die Seitenlinie lebenslang erhalten. Die Befruchtung erfolgt bei den meisten Arten innerlich, das Weibchen fängt mit der Kloake die Schleimsäcke mit Spermatozoen (Spermatophoren) ein.
Einige beobachteten den Fortpflanzungsprozess im Larvenstadium - Neotenie. Dieses Phänomen ist am bekanntesten für den Axolotl, eine nordamerikanische Amphibie, die fortpflanzungsfähig ist, aber eigentlich das Larvenstadium des Ambystoma ist.
Bestellen Sie schwanzlose Amphibien (Anura). Schwanzlos - die am besten organisierte und zahlreichste Gruppe von Amphibien mit derzeit 2900 Arten. Bei aller Artenvielfalt äußere Struktur eher eintönig, was mit Bewegung durch Springen einhergeht. Die Körperform ist verkürzt, keilförmig. Die Hinterbeine sind verlängert, in ihnen erscheint ein neuer Abschnitt, der „sekundäre Unterschenkel“, der durch Verlängerung der beiden Fußwurzelknochen gebildet wird. Es sollte beachtet werden, dass die paarigen Knochen im Unterschenkel und Unterarm für eine bessere Unterstützung bei der Bewegung an Land verschmolzen sind.
Die Abteilung ist in 19 Familien unterteilt, von denen die bekanntesten sind: die Familie der Rundzungen (Kröten), die Familie der Pipaceae (Spornfrosch, Surinamische Pipa), die Familie der Knoblauchkröte (gemeine Knoblauchkröte), die Familie der Echten Kröten (graue und Wechselkröten), die Froschfamilie (Gemeiner Laubfrosch), eine Familie echter Frösche (See- und Teichfrösche).
Schlüsselbegriffe und Konzepte
1. Spiralventil. 2. Rumpfnieren. 3. Urethel. 4. Wolfskanal. 5. Müllers Kanal. 6. Mittelohr. 7. Eustachische Röhre. 8 Neotenie. 9. Ripidistie. 10. Ichthyostegiden. 11. Stegozephalie.
Wesentliche Überprüfungsfragen
In welche Ordnungen werden moderne Amphibien eingeteilt?
Welche Eigenschaften haben Amphibien im Zusammenhang mit der Landung erworben?
Welche Abteilungen werden in den vorderen und hinteren Gliedmaßen unterschieden?
Welche Abteilungen werden in der Wirbelsäule unterschieden?
Was sind der Schulter- und Beckengürtel der Gliedmaßen?
Welche großen Verdauungsdrüsen münden in den Zwölffingerdarm?
Wie heißt der letzte Darmabschnitt, in den Dickdarm, Ausscheidungs- und Fortpflanzungssystem münden?
Was sind die Atmungsorgane von Larven und erwachsenen Amphibien?
Wie viele Blutkreisläufe und wie viele Kammern gibt es im Herzen erwachsener Amphibien?
Wie viele Blutkreisläufe und wie viele Kammern hat das Herz einer Kaulquappe?
Welches Blut gelangt in den rechten und linken Vorhof?
Welche Organe entstehen Ausscheidungssystem Amphibien?
Was sind die Nieren erwachsener Amphibien?
In welcher Form werden die Produkte des Stickstoffstoffwechsels bei erwachsenen Amphibien ausgeschieden?
In welcher Form werden die Produkte des Stickstoffstoffwechsels in Amphibienlarven ausgeschieden?
Wie unterscheiden sich die Hör- und Sehorgane von Amphibien von denen von Fischen?
Welche Eigenschaften ermöglichten es Fischen, den terrestrischen Lebensraum zu erobern und sich in Amphibien zu verwandeln?
Phylogenie der Amphibien.
Alle Wirbeltiere haben spezialisierte Atmungsorgane (mit Ausnahme einiger Amphibien, die sie zum zweiten Mal verloren haben). Diese Organe entwickeln sich in der Regel aus Auswüchsen des Darmschlauches (oben wurde auf die phylogenetische Beziehung zwischen Atmungs- und Ernährungsfunktion hingewiesen).
Das allgemeine Prinzip der Funktionsweise von Atmungssystemen besteht darin, zu schaffen dynamischer Kontakt von Blut mit einem gasförmigen Medium, in Kombination des Blutflusses in den Gefäßen der Atmungsorgane (deren Perfusion) mit dem Pumpen von Wasser oder Luft durch diese Organe (deren Belüftung). Eine Ausnahme bildet die Hautatmung von Amphibien, die kein spezielles Beatmungsgerät benötigt.
Wasser atmen.Die Wasseratmung mit Hilfe von Kiemen ist den meisten Fischen eigen. Diese Tiere ziehen Wasser in die Mundhöhle, wonach das Wasser durch die Kiemenschlitze herausgedrückt wird. Verwendet, um Sauerstoff zu extrahieren Gegenstromprinzip(Abb. 10.4). Blut in den Kiemenkapillaren fließt in Richtung des Wassers, das die Kiemenblätter wäscht, was durch die Bewegung der Fische im Weltraum erleichtert wird. Die hohe Effizienz der Kiemenatmung soll den großen Energieaufwand für die Arbeit der Muskeln kompensieren, die die Kiemen mit Wasser belüften. Dieser Muskel wird von einem separaten gesteuert Atmungszentrum
|
Reis. 10.4. Kiemenbögen (vier) bei Fischen. (A), ein Fragment von zwei Kiemenbögen (B), ein Abschnitt eines Kiemenfadens mit drei Kiemenfäden auf jeder Seite (C) und ein Abschnitt von knöchernen Fischkiemen, in denen Blut und Wasser in entgegengesetzte Richtungen fließen (D) 1 - Kiemendeckel, 2 - Konstriktormuskel, 3 - Kiemenfäden, 4 - Kiemenhöhle, 5 - Blutgefäße, 6 - Mundhöhle, 7 - Kiemenskelett, 8 - Platten, 9 - Querschnitt durch das Kiemenfilament, 10 - Septum, 11 - Septumkanal, 12 - Rand der Platte. |
vMedulla oblongata, die sich in ihrer Organisation dem Atmungszentrum von Säugetieren nähert. Entdeckung der rhythmischen Aktivität von Strukturen im isolierten Hirnstamm eines Goldfisches ( Carassius Auratus Auratus) war ein wichtiger Meilenstein in der Erkenntnis Automatismus des Atemzentrums.
Aufgrund der hohen Löslichkeit von CO 2 in Wasser wird es leicht freigesetzt, sodass die Arbeit des Atmungsapparates der Fische hauptsächlich dazu dient, den Körper mit Sauerstoff zu versorgen. Signale über den O2-Gehalt im Blut, die von den Chemorezeptoren der Kiemengefäße kommen, regulieren nicht nur die Aktivität des Atemzentrums des Fisches, sondern veranlassen ihn auch, Zonen des Reservoirs mit physiologisch optimaler Sauerstoffversorgung zu wählen. Diese Verhaltensreaktion des aktiven Auswählens einer bevorzugten Atmungsumgebung wird als bezeichnet Gas bevorzugt. Einige Fische (Aale) nehmen im Wasser gelösten Sauerstoff hauptsächlich nicht über die Kiemen auf, sondern nutzen dessen Diffusion durch die Haut.
Übergang von der Wasser- zur Luftatmung. Viele Fische, die sporadisch an die Oberfläche von Gewässern aufsteigen oder sogar an Land gehen und Luft schlucken, ergänzen die Wasseratmung mit Luft, wofür sie die reich vaskularisierten Wände der Mundhöhle, des Rachens, des Magens, des Darms oder eines speziellen Derivats davon nutzen das Darmrohr - Schwimmblase.
Lungenfisch. atmen Sie bereits mit Hilfe eines neuen spezialisierten Organs - Lunge Dies ist ein gefalteter Sack (einzeln oder paarig), der aus dem ventralen Vorsprung des Pharynx stammt. Einige Arten von zweibeinigen Fischen ( Lepidosireniformes) leben in tropischen Gewässern, schlecht belüftet und auch übersäuert. Ohne Atemluft kommen sie nicht aus.
|
|
|
Reis. 10.5 Das Funktionsprinzip der Atmungssysteme von Wirbeltieren 1 - Blut, 2 - Barriere, 3 - Umgebung, 4 - Atmungsorgane, 5 - Diffusion; Pi, Re - Partialdrücke von O 2 in der eingeatmeten und ausgeatmeten Luft (Wasser); Pa, Pv- das gleiche in arteriellem und venösem Blut. |
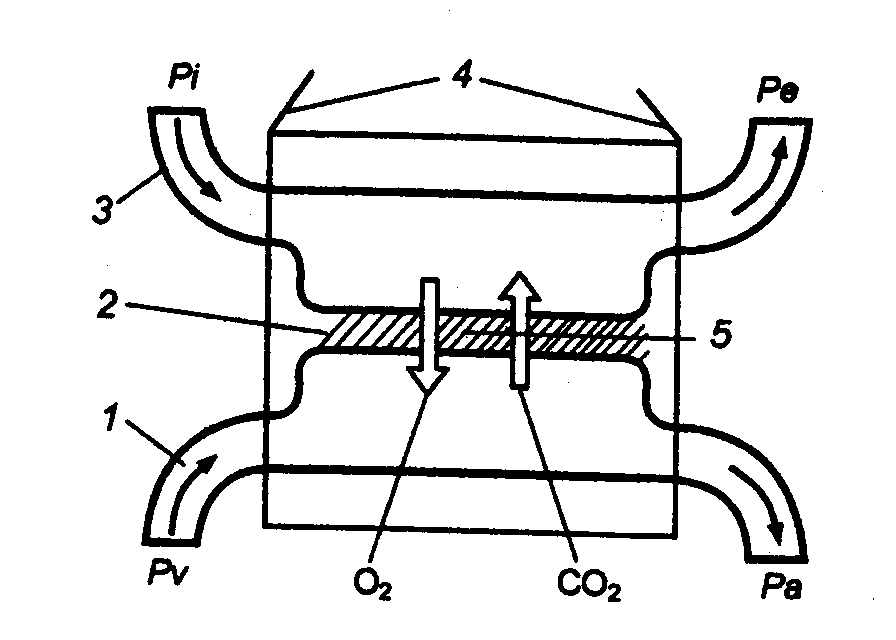
Eine solche Veränderung der Bedingungen trug offensichtlich zum Massenübergang von aquatischen Wirbeltieren (es waren Lappenflossenfische - die Vorfahren der Amphibien) zur Luftatmung in der Devonzeit (vor etwa 400 Millionen Jahren) bei. Es sollte berücksichtigt werden, dass der Partialdruck von 02 in der Atmosphäre damals dem heutigen nahe kam. Und wenn 1 Liter mit Luft gesättigtes Wasser bei einer Temperatur von 0 ° C nur 0,014 g 02 enthält (bei 30 ° C halbiert sich diese Menge), dann beträgt sein Gehalt in 1 Liter Luft 0,28 g, d. H. in 20-mal mehr . Zudem ist die Dichte von Luft mehr als 700-mal geringer als die von Wasser, sodass ihre Belüftung durch die Atmungsorgane entsprechend weniger Energieaufwand erfordert. Die meisten Amphibien (zumindest ihre erwachsenen Formen) haben auf Luftatmung umgestellt, obwohl die Lungen der Amphibien noch schwach entwickelt sind: Die Gasaustauschfläche ist klein, die venösen und arteriellen Blutströme im Kreislaufsystem werden nicht vollständig verdünnt. Ein Teil des aufgenommenen O 2 und der größte Teil des emittierten CO 2 diffundieren nicht durch die Lunge, sondern durch die angefeuchtete Haut, deren Vaskularisation sich entsprechend der Intensität des Hautgasaustausches verändert (Abb. 10.5).
Atembewegungen Amphibie(wie Reptilien) sind charakteristisch - darin unterscheiden sie sich sowohl von Fischen als auch von Säugetieren - lange Stopps, Apnoe, dabei wird nur der Mund-Rachen-Raum bei geschlossener Glottis belüftet, die die Lunge von der Atmosphäre trennt. Nur von Zeit zu Zeit gibt es koordinierte Kontraktionen der Mund- und Kehlkopfmuskulatur, die Luft in die Lunge drücken und zurücksaugen. Diese Bewegungen werden von den Nervenzentren des Hirnstamms entsprechend den Impulsen der Dehnungsrezeptoren der Lunge sowie der arteriellen Chemorezeptoren gesteuert, die durch Sauerstoffmangel im Blut angeregt werden. Darüber hinaus weisen viele Amphibien eine Verhaltensregulation der Atmung auf, eine Gaspräferenz: Je nach Bedingungen wählen sie entweder eine aquatische oder eine Luftumgebung.
Terrestrisch Reptilien im Zusammenhang mit der Entwicklung der verhornenden Hülle wird die Wasser- und Hautatmung unmöglich (mit Ausnahme von Eidechsen). Dieses Stadium in der Evolution der Atmung markiert den endgültigen Übergang vom Gasaustausch in einer feuchten Umgebung zur "trockenen" Ventilation, deren Mechanismus bereits fast derselbe ist wie bei Säugetieren. Die Gasaustauschfläche der Lunge wird durch die Bildung kleiner Zellen deutlich vergrößert, das Blut eines bereits isolierten kleinen Kreises fließt durch ein dichtes Netz ihrer Gefäße. Gleichzeitig behalten Reptilien immer noch die intermittierende Art der Belüftung der Lunge bei, die jedoch mit Luft gefüllt wird, indem sie nicht forciert, sondern eingesaugt wird.
Stoffwechsel wechselwarme (poikilotherme) Wirbeltiere noch relativ gering, was der Grund für die relativ geringe "Kapazität" ihres Atemapparates ist. Einigen Forschern zufolge war dies der Grund für die Unfähigkeit der heute ausgestorbenen großen Reptilien, sich an die dramatischen Veränderungen des Lebensraums am Ende des Mesozoikums anzupassen.
Deutlich intensivere und stabilere Stoffwechselvorgänge in Endothermen (homöotherme Tiere) - Säugetiere und Vögel. Die Gasaustauschfläche und die Durchblutung der Lunge nehmen weiter zu. Im Gegensatz zu Amphibien und Reptilien eine neu gebildete rhythmisch, ohne Pausen, Art der Lungenbeatmung.Ändert den Mechanismus der Regulation der Atmung erheblich, da das Ganze CO2 bei diesen Tieren wird es praktisch nur durch die Lunge entfernt, und seine Produktion wird durch die Intensivierung oxidativer Prozesse sehr intensiv. Es gibt eine Schwierigkeit bei der Bekämpfung der Ansammlung von Kohlensäure in Geweben und Blut. Daher steigt bei Tieren, die ausschließlich mit der Lunge atmen, der arterielle Blutdruck zwar an, gleichzeitig steigt aber auch der Pco 2 deutlich an (Abb. 10.6) und auch die Leistungsfähigkeit von Puffersystemen nimmt zu.
|
|
|
Reis. 10.6 Atemgasdruck im arteriellen Blut (PaO 2 Paco 2 ) bei verschiedene Klassen Wirbeltiere Die Kiemenatmung von Fischen versorgt sie mit Sauerstoff; R co 2 ihr Blut ist niedrig, weil co 2 diffundiert leicht durch die Haut des Körpers in das umgebende Wasser. Atemverhalten und Kiemenatmung bei Fischen werden durch Sauerstoff reguliert. Für diese Fische das bestimmte Bedingungen Die Luftatmung (wie auch bei Reptilien und Säugetieren) nutzt nicht nur die Aufnahme von O 2 , sondern auch die Rückführung co 2 in die Luft erfolgt nur über die Lunge. Deshalb R co 2 Blut bei luftatmenden Tieren ist viel höher als bei wasseratmenden Tieren, und dieser Parameter wird zum Hauptregulator der Atmung. Allerdings R o 2 Blut in Wasser- und Luftatmer unterscheiden sich geringfügig. |

Aufgrund dieser Umstände geht die Rolle des Hauptregulators der Atmung von O 2 auf CO 2 und die mit seinem Transport verbundenen H + -Ionen über, und die zentralen (medullären) Ionen übernehmen die führende Rolle bei der Stimulierung der rhythmischen Aktivität des Atmungszentrums statt arterieller Chemorezeptoren. chemosensitive Bereiche. Alle diese Funktionen werden in den entsprechenden Abschnitten des Kapitels behandelt. Gleichzeitig unterscheidet sich die Atmung von Vögeln so sehr von der Atmung von Säugetieren, dass wir uns gesondert damit befassen sollten.
Atem der Vögel.Die Lungen von Vögeln, die eine schwammartige Struktur haben, unterscheiden sich zum einen dadurch, dass der Gasaustausch zwischen Luft und Blut in den sogenannten Luftkapillaren, "zylindrischen Alveolen" (in Analogie zu den sackförmigen Alveolen von Säugetieren) stattfindet - und zum anderen , im Bronchialflusssystem . Dieses System ist so konzipiert, dass die Lunge mit Hilfe mehrerer Luftsäcke werden sowohl beim Ein- als auch beim Ausatmen mit Frischluft belüftet (siehe Abb. 10.7). Diese Art der Atmung verleiht dem pulmonalen Gasaustausch des Vogels eine hohe Effizienz, die notwendig ist, um den intensiven Energieverbrauch, insbesondere im Flug, zu decken. Die Atembewegungen der Brustwand werden hauptsächlich durch Kontraktionen der Brust- und Bauchmuskulatur erzeugt. Im Flug trägt die Belüftung der Lunge (die neben dem Gasaustausch an der Abgabe überschüssiger Wärme beteiligt ist) zur Bewegung der Flügel bei.
Das Atmungszentrum des Vogels befindet sich in der Medulla oblongata. Seine Aktivität wird wie bei Säugetieren durch Impulse von medullären Chemorezeptoren gesteuert, die für CO 2 und H + -Ionen empfindlich sind, sowie von arteriellen Chemorezeptoren, die auch auf O 2 -Mangel ansprechen, der für die Regulierung der Atmung während des Fluges wichtig ist auf großen Höhen. In den Atemwegen des Vogels befinden sich Mechanorezeptoren, deren Stimulation an der Regulierung der Atemfrequenz und -tiefe beteiligt ist. Es gibt auch spezielle Chemorezeptoren, deren Impulsation mit abnehmender intrapulmonaler Konzentration zunimmt CO2 . Die Stimulation dieser Rezeptoren hemmt die Aktivität des Atmungszentrums und reduziert dadurch die Lungenventilation, wodurch eine übermäßige CO 2 -Auswaschung aus dem Körper verhindert wird, die unter Flugbedingungen auftreten könnte.