Bildung von Geweben des Stammes der Sekundärstruktur
Die Sekundärstruktur des Stängels ist charakteristisch für einjährige und mehrjährige krautige, holzige Dikotyledonen sowie Gymnospermen. Bei zweikeimblättrige Pflanzen Die Primärstruktur ist sehr kurzlebig und mit Beginn der Aktivität des Kambiums bildet sich eine Sekundärstruktur. Abhängig von der Verlegung des Prokambiums werden verschiedene Arten der Sekundärstruktur des Stängels gebildet. Werden die Stränge des Prokambiums durch breite Parenchymreihen getrennt, so entsteht eine Balkenstruktur, werden sie so zusammengeführt, dass sie zu einem Zylinder verschmelzen, entsteht eine Nichtbalkenstruktur.
Reis. 3.24. Balkenstruktur des Stängels einer zweikeimblättrigen Pflanze: A - Klee: 1 - Epidermis; 2 - Chlorenchym; 3 - Sklerenchym perizyklischen Ursprungs; 4 - Phloem; 5 - gebündeltes Kambium; 6 - Xylem; 7 - interfaszikuläres Kambium
Balkenstruktur des Stiels kommt in Pflanzen wie Klee, Erbsen, Ranunkel, Dill vor (Abb. 3.24). Ihre Prokambialstränge sind in einem Kreis entlang der Peripherie des Zentralzylinders verlegt. Jeder Prokambialstrang verwandelt sich in ein Kollateralbündel, das aus primärem Phloem und primärem Xylem besteht. Später wird zwischen Phloem und Xylem das Kambium aus dem Prokambium gebildet, das die Elemente des sekundären Phloems und des sekundären Xylems bildet. Phloem wird an der Peripherie des Organs abgelagert, und Xylem wird in der Mitte abgelagert, und noch mehr Xylem wird abgelagert. Das primäre Phloem und das Xylem bleiben an der Peripherie des Bündels, während die sekundären Elemente an das Kambium angrenzen. Die Stängel zweikeimblättriger Pflanzen zeichnen sich durch die Bildung offener Kollateral- oder Bikollateralbündel aus (Abb. 17, siehe Farbakzente).
Auch die Stängel zweikeimblättriger Pflanzen zeichnen sich durch Differenzierung aus primärer Kortex, Dazu gehören: Kollenchym (Ecke (Abb. 18, siehe Farbe inkl.) oder Lamellen), Chlorophyll tragendes Parenchym und die innere Schicht - Endoderm. Stärke reichert sich im Endoderm an; solch stärkehaltige Vagina Theaterstücke wichtige Rolle in der geotropen Reaktion der Stängel. Am Rand der Primärkruste befindet sich der zentrale Axialzylinder perizyklisches Sklerenchym- ein durchgehender Ring oder Bereiche in Form von Halbbögen über dem Phloem. Der Kern des Stammes wird durch das Parenchym ausgedrückt und repräsentiert. Manchmal wird ein Teil des Kerns unter Bildung eines Hohlraums zerstört (siehe Abb. 3.24).
Struktur ohne Träger charakteristisch für Holzgewächse(Linde) (Abb. 19, siehe Spalte inkl.) und viele Kräuter (Flachs). Im Wachstumskegel verschmelzen die Prokambialstränge und bilden einen durchgehenden Zylinder, der im Querschnitt als Ring sichtbar ist. Der Ring des Prokambiums bildet nach außen den Ring des primären Phloems und nach innen den Ring des primären Xylems, zwischen dem der Ring des Kambiums liegt. Kambiumzellen teilen sich (parallel zur Oberfläche des Organs) und legen einen Ring aus sekundärem Phloem nach außen und einen Ring aus sekundärem Xylem nach innen im Verhältnis 1:20 ab. Wir betrachten die Nichtbalkenstruktur am Beispiel eines mehrjährigen verholzenden Lindenstamms (Abb. 3.25).
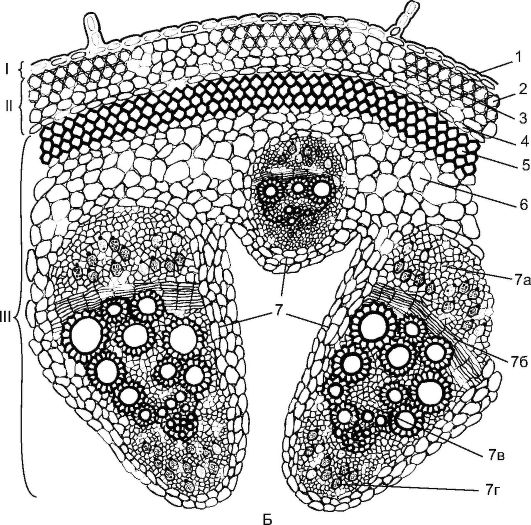
Reis. 3.24.(Fortsetzung) B – Kürbis: I – Hautgewebe; II – Primärrinde; III - zentraler Axialzylinder; 1 - Epidermis; 2 - Eckkollenchym; 3 - Chlorenchym; 4 - Endoderm; 5 - Sklerenchym; 6 - Hauptparenchym; 7 – bikollaterales vaskuläres Faserbündel: 7a – Phloem; 7b - Kambium; 7c - Xylem; 7d – inneres Phloem
Ein junger Lindenspross, der sich im Frühjahr aus einer Knospe bildet, ist mit Epidermis bedeckt. Als Rinde werden alle dem Kambium vorgelagerten Gewebe bezeichnet. Die Rinde ist primär und sekundär. Primärer Kortex Es besteht aus einem lamellaren Kollenchym, das sich unmittelbar unter der Epidermis in einem durchgehenden Ring befindet, einem chlorophyllhaltigen Parenchym und einer einreihigen stärkehaltigen Hülle. Diese Schicht enthält Körner „geschützter“ Stärke, die die Pflanze nicht verbraucht. Es wird angenommen, dass diese Stärke an der Aufrechterhaltung des Pflanzengleichgewichts beteiligt ist.
Der zentrale Axialzylinder der Linde beginnt mit einem perizyklischen Sklerenchym oberhalb des Phloems. Durch die Aktivität des Kambiums sekundärer Kortex(vom Kambium bis zum Periderm), dargestellt durch sekundäres Phloem, Kernstrahlen und Parenchym des sekundären Kortex. Die Rinde der Linde wird geerntet, indem man sie zum Kambium entfernt. Dies ist besonders einfach im Frühjahr, wenn sich die Kambiumzellen aktiv teilen. Früher wurde Lindenrinde (Bast) zum Weben von Bastschuhen, zur Herstellung von Kisten, Waschlappen usw. verwendet.
Das trapezförmige Phloem ist durch dreieckige primäre Kernstrahlen unterteilt, die das Holz bis zum Kern durchdringen. Die Zusammensetzung des Phloems der Linde ist heterogen. Es enthält verholzte Bastfasern, aus denen Hartbast besteht, und Weichbast wird durch Siebröhren mit Satellitenzellen und Bastparenchym dargestellt. Der Bast verliert in der Regel nach einem Jahr seine Fähigkeit, organische Stoffe zu leiten, und erneuert sich durch die Aktivität des Kambiums mit neuen Schichten.
Das Kambium bildet auch sekundäre Kernstrahlen, diese erreichen den Kern jedoch nicht und gehen im Sekundärholz verloren. Die Kernstrahlen dienen der Bewegung von Wasser und organische Substanz in radialer Richtung. In den Parenchymzellen der Kernstrahlen werden bis zum Herbst Reservenährstoffe (Stärke, Öle) eingelagert, die im Frühjahr für das Wachstum junger Triebe verbraucht werden.
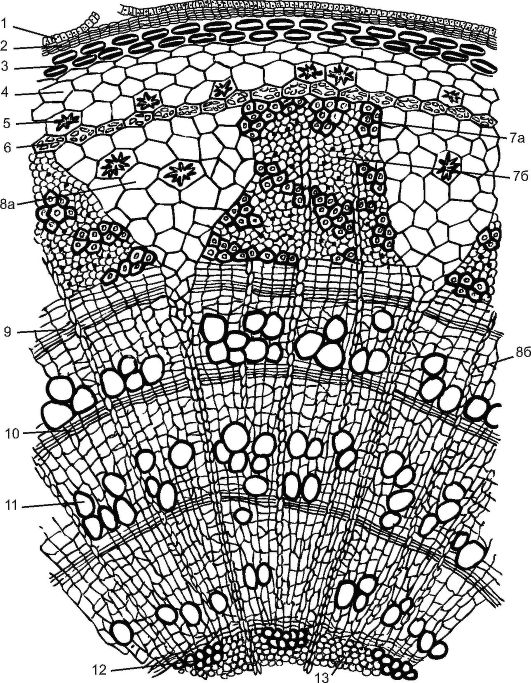
Reis. 3.25. Querschnitt eines drei Jahre alten Lindenzweigs: 1 - Reste der Epidermis; 2 - Kork; 3 - lamelläres Kollenchym; 4 - Chlorenchym; 5 - Drusen; 6 - Endoderm; 7 – Phloem: 7a – harter Bast (Bastfasern); 7b – Weichbast – (Siebröhren mit Begleitzellen und Bastparenchym); 8a – primärer Kernträger; 8b – sekundärer Kernträger; 9 - Kambium; 10 - Herbstholz; 11 - Federholz; 12 - primäres Xylem; 13 - Kernparenchym
Bereits im Sommer wird Phellogen unter die Epidermis gelegt und ein sekundäres Hautgewebe, das Periderm, gebildet. Im Herbst sterben mit der Bildung des Periderms die Zellen der Epidermis ab, ihre Überreste bleiben jedoch 2-3 Jahre bestehen. Die Schichtung des mehrjährigen Periderms bildet eine Kruste.
Die vom Kambium in Gehölzen gebildete Xylemschicht ist viel breiter als die Phloemschicht. Holz funktioniert mehrere Jahre. Abgestorbene Holzzellen sind nicht an der Stoffleitung beteiligt, können aber das enorme Gewicht der Pflanzenkrone tragen.
Die Zusammensetzung von Holz ist heterogen und umfasst: Tracheiden(Abb. 20, siehe Farbe inkl.), Luftröhre, Holzparenchym Und libriform. Holz zeichnet sich aus durch Jahresringe. Im zeitigen Frühjahr, wenn in der Pflanze ein aktiver Saftfluss stattfindet, bildet das Kambium im Xylem weitlumige und dünnwandige leitende Elemente – Gefäße und Tracheiden. Mit dem Herannahen des Herbstes, wenn diese Prozesse aufhören und die Aktivität des Kambiums nachlässt, erscheinen englumige dickwandige Gefäße, Tracheiden und Holzfasern. Dadurch entsteht ein Jahreswachstum bzw. ein Jahresring (von einer Quelle zur anderen), der im Querschnitt deutlich sichtbar ist. Anhand der Jahresringe können Sie das Alter der Pflanze bestimmen (siehe Abb. 3.25).
Merkmale der Struktur des Stammes von Dikotyledonen:
1) Dickenwachstum des Stängels (aufgrund der Aktivität des Kambiums);
2) gut differenzierter primärer Kortex (Collenchym, Chlorophyll-tragendes Parenchym, Stärke-tragendes Endoderm);
3) bikollaterale und kollaterale Bündel nur offener Art (mit Kambium);
4) Gefäßfaserbündel liegen ringförmig oder verschmelzen (Nicht-Bündel-Struktur);
5) das Vorhandensein eines Kerns;
6) Gehölze zeichnen sich durch das Vorhandensein von Wachstumsringen im Xylem aus.
Strukturmerkmale dikotyler Rhizome. Das Deckgewebe zweikeimblättriger Rhizome kann die Epidermis sein, während bei mehrjährigen Rhizomen die Epidermis durch das Periderm ersetzt wird. Der primäre Kortex wird durch Speicherparenchym und Endoderm mit Casparian-Flecken dargestellt. Darüber hinaus nähert sich die Breite der Primärkruste der Breite des Zentralzylinders an. Die Struktur des zentralen Axialzylinders, der Gefäßfaserbündel und ihre Lage darin weisen die gleichen Merkmale auf wie bei oberirdischen Stängeln.
Abgerundet (zum Beispiel: Sonnenblume)
Tetraeder
dreieckig
Polyedrisch (gerippt) (zum Beispiel: Kürbis)
Geflügelt (mit ledrigen Auswüchsen)
Stammbildung
Der Stamm wird wie die Wurzel aus dem primären Apikalmeristem gebildet. Im Gegensatz zur Wurzel ist der Wachstumskegel geschützt Umfeld junge, sich entwickelnde Blättchen. Wie in der Wurzel teilen sich Meristemzellen durch Mitose und spezialisieren sich auf primäres permanentes Gewebe, aber im Gegensatz zur Wurzel gibt es im Stamm keine klar definierten Wachstumszonen. Die Zellspezialisierung erfolgt sehr schnell und genau wie die Wurzel spezialisieren sich diese Zellen auf drei gleiche Bereiche.
Von der Peripherie geht 1 Zellschicht in die primäre über Hautgewebe; im primären Kortex; Im Zentrum befindet sich ein zentraler Axialzylinder. Im Stamm nimmt der Zentralzylinder im Gegensatz zur Wurzel ein Vielfaches mehr Platz ein als die Primärrinde. 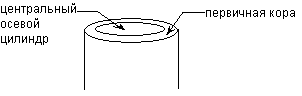
Primäres Hautgewebe Stammepidermis, 1 Schicht, mit Stomata, mit Haaren. Seine Funktion besteht darin, darunter liegende Strukturen zu schützen.
Primärer Kortex Der Stängel ist für die Photosynthese verantwortlich und sorgt für mechanische Stabilität (biegt nicht ab). Definiert einen Stamm im Raum. Es besteht aus:
1) Chlorenchym (von 1 bis vielen Schichten dieser Zellen)
2) Kollenchym (aus einer oder mehreren Schichten). Selten bei Monokotyledonen zu sehen.
3) Endoderm
Das Endoderm des Stängels ist eine Schicht lebender Zellen mit Zellulosemembranen, die Stärkekörner enthalten. Sie sind keine Speichersubstanz, sie helfen bei der Navigation im Weltraum und werden daher als Endodermis des Stängels bezeichnet stärkehaltigVagina. Darüber hinaus können sich bei verschiedenen Pflanzen in der Primärrinde Sekretbehälter bilden, es finden sich kristalltragende Zellen, das ist der plastische Teil des Stängels.
Zentralachszylinder es beginnt mit einem perizyklischen Sklerenchym. Der Perizykel im Stängel degeneriert zum Sklerenchym und teilweise zum Parenchym. Perizyklisches Sklerenchym kann im Querschnitt in Form eines durchgehenden Rings oder in Form einzelner Stränge vorliegen.
Hinter dem Perizykel befinden sich meist kollotyrale Leitbündel; selten bikollotural, umgeben von Zellen des Hauptparenchyms.
Die Anordnung der Leitbündel ist bei Monokotyledonen und Dikotyledonen unterschiedlich.
- bei Monokotyledonen ist es diffus (verstreut) lokalisiert
- - in Dikotyledonen im Kreis
Eine solche unterschiedliche Anordnung der Leitbündel hängt mit der Art der Kombination der Leitbündel von Blatt und Stängel zusammen.
Bei Dikotyledonen sind die Leitblattbündel an ihrem Knoten mit dem Stamm verbunden. ![]()
Bei einkeimblättrigen Pflanzen dringen die Blattbündel in den zentralen Zylinder ein, biegen sich und verbinden sich mit Stammbündeln, die durch 1–2 Knoten verlaufen. 
Bei Dikotyledonen dringen 2-3 Blattbündel von einem Blatt in den Stängel ein, bei Monokotyledonen sind es aufgrund der Paralleladerung viele.
Bei einkeimblättrige Pflanzen alle leitfähigen Bündel geschlossen, d.h. haben kein Kambium, während sie in Dikotyledonen vorkommen offen Bündel, d.h. hat ein Kambium.
Die Dicke des Stängels einkeimblättriger Pflanzen bleibt über die gesamte Länge gleich, während bei zweikeimblättrigen Pflanzen der obere Teil des Stängels etwas geringer ist als der untere Teil.
Abhängig von Umweltbedingungen, in dem dieser oder jener Pflanzentyp gebildet wird, werden verschiedene Merkmale der Struktur seiner Organe, einschließlich des Stängels, geschaffen.
Darüber hinaus übernimmt der Stamm in manchen Fällen die Funktionen anderer Organe. Dabei sind Veränderungen in der Stammstruktur besonders bedeutsam und werden meist als Anomalien eingestuft. Kriechpflanzen zeichnen sich durch eine besondere Struktur des Stängels aus. Besonders reich sind Lianen in den Tropen vertreten, wo sie am vielfältigsten und kräftigsten entwickelt sind. Aber auch in gemäßigten Breiten wachsen genügend Reben. Hopfen, Clematis, Efeu, Weinrebe, Erbsen, Bohnen, Glyzinien, Ackerwinde usw. sind echte Schlingpflanzen, nicht so luxuriös wie in den Tropen, weisen aber dennoch Merkmale in der Stängelstruktur auf, die allen Schlingpflanzen eigen sind.
Das charakteristischste Merkmal der Rebenstammstruktur ist die ziemlich ausgeprägte Dissektion des Leitungssystems und die starke Entwicklung der parenchymalen Markstrahlen. Auch in Lianen mit gut definiertem holziger Stamm(Weinreben-)Kernstrahlen bestehen aus einem Parenchym, das sich kaum vom Parenchym der Rinde und des Kerns unterscheidet, und diese Kernstrahlen sind sehr hoch, erstrecken sich in großer Entfernung über die Länge des Stiels und ähneln langen Bändern, die vertikal entlang des Stiels angeordnet sind und die Masse des zentralen Zylinders in separate Sektoren zerlegen. Bei typischen Gehölzen wirken die Kernstrahlen aufgrund der unbedeutenden Höhe an tangentialen Holzabschnitten wie kurze spindelartige Zellhaufen. Bei tropischen Reben ist die Teilung des Mittelzylinders des Stängels noch schärfer. Bei einigen tropischen Reben beginnen sich an verschiedenen Stellen im Holz des Zentralzylinders verstreute Parenchymzellen intensiv zu teilen und zu wachsen, wodurch einzelne Abschnitte des Gefäßsystems auseinandergedrückt werden. Dadurch zeichnen sich die alten Stängel solcher Reben durch eine sehr unregelmäßige und skurrile Struktur aus. Im Allgemeinen ähnelt die Struktur des Stammes von Gehölzen der Struktur des Stammes einiger krautiger Pflanzen.
Bei einer Reihe von Pflanzen, die in den Wüstengebieten Afrikas und Amerikas wachsen, wo es in der Regel nur sehr selten und dann nur für kurze Zeit im Jahr regnet, weisen die Form und Struktur des Stängels besondere Besonderheiten auf. Bei solchen Pflanzen sind die Stängel fleischig, grün, nehmen eine andere Form an – eine Kugel, einen Teller oder sind mit breiten Lamellenrippen ausgestattet. Diese Stängel dienen als Blätter. Die Blätter sind oft sehr klein und sehen manchmal wie trockene Stacheln aus. Als Pflanzen werden Pflanzen mit fleischigen Stängeln und reduzierten Blättern bezeichnet Stamm Sukkulenten. Darunter sind verschiedene Arten von Kaktusfeigen, Kakteen und Wolfsmilch.
Die Reduzierung der Blattspreiten auf trockene Stacheln führt zu einer erheblichen Verringerung der Verdunstungsoberfläche der gesamten Pflanze, die in ihrem oberirdischen Teil vollständig aus einem eigenartig veränderten Stängel besteht. Der Stängel selbst verwandelt sich in ein Organ, das zur Ansammlung von Wasserreserven geeignet ist. Dadurch kann die Pflanze in Regenperioden Feuchtigkeit speichern und in Trockenperioden langsam verbrauchen.
In der Struktur des Hautgewebes der meisten Arten von Sukkulentenstämmen gibt es Anpassungen, um möglicherweise den Wasserverlust der Pflanze durch Transpiration zu reduzieren. In diesem Fall sollte auch das System der leitfähigen Gewebe stark reduziert werden. Das mechanische Gewebe in den Sukkulentenstängeln wird überflüssig und ist fast nicht mehr vorhanden.
Der Zellsaft von Gewebezellen sukkulenter Stängel besteht aus Lösungen verschiedener Säuren, Zucker und Schleimstoffen. Diese Stoffe tragen dazu bei, den Wasserrückfluss zur Pflanze zu reduzieren. Das Gewebe saftiger Stängel ist sehr homogen und besteht hauptsächlich aus dünnwandigem Parenchym, das von unterentwickelten Gefäßbündeln durchzogen ist: In jedem Bündel befinden sich nur wenige Gefäße, sie sind alle eng hohl und von primitiver Struktur. Die schwache Entwicklung der Leitbündel steht durchaus im Einklang mit der unbedeutenden Entwicklung der Blätter, da das Blatt in seiner Entwicklung die Bildung von Leitbündeln des Stängels stimuliert. In typischen, gut ausgeprägten Sukkulentenstämmen wird aufgrund der schwachen Kambialaktivität wenig sekundäres Xylem gebildet, das Gefäßsystem besteht hauptsächlich aus Primärelementen, die im Gefäßmeristem entstanden sind. Daher ist die Ausbildung eines schwachen Leitungssystems in den Stängeln von Sukkulenten sowohl physiologisch als auch entwicklungsgeschichtlich durchaus verständlich.
Unter den Sukkulenten gibt es natürlich keine holzigen Formen, alle sind in ihrer Struktur krautige Pflanzen.
Pflanzen der sogenannten sklerenchymaler Typ, wächst auch an trockenen Orten, normale Blätter und Stängel sind entwickelt, das Leitsystem besteht aus kräftig entwickelten vaskulären Faserbündeln oder deren Fusionsprodukten. Die Gefäßelemente in solchen Bündeln sind breitbandig, zahlreich und besser organisiert als bei Sukkulenten. Sekundäre Elemente kommen in Gehölzen dieser Gruppe deutlich zum Ausdruck, und in den Stängeln krautiger Pflanzen ist die Verholzung der Zellmembranen des Hauptparenchyms ausgeprägt. Das mechanische System ist hoch entwickelt, seine anatomischen Elemente sind dickwandig.
Die Stämme der im Wasser wachsenden Pflanzen – der sogenannten Wasserpflanzen – mit einer relativ großen Formenvielfalt haben ein gemeinsames Strukturmerkmal: Sie verfügen über ein gut entwickeltes System von Interzellulargängen und Interzellularräumen. Interzelluläre Räume in Form von mehr oder weniger großen Hohlräumen bilden ein System von Luftkanälen, die sowohl den Kern als auch die Rinde des Stängels durchdringen. Gleichzeitig ist der Kern besonders stark entwickelt und nimmt einen erheblichen Anteil des Gesamtdurchmessers des Stiels ein. Bei Wasserpflanzen, insbesondere bei Vertretern der Klasse der Monokotyledonen, überwiegt manchmal das mechanische System gegenüber dem Leitungssystem. In Bündeln gesammelte Gruppen vaskulärer anatomischer Elemente sind normalerweise relativ klein, während Anordnungen, die aus mechanischen Elementen bestehen und hauptsächlich entlang der Peripherie des Stamms konzentriert sind, im Vergleich zu Gefäßbündeln eine große Leistung erreichen.
Noch stärker weicht die Struktur der unterirdischen Stängel, die die Funktion von Vorratsbehältern erfüllen - Rhizome und Knollen -, vom normalen Typ ab. Das Rhizom ist ein mehrjähriger Trieb und erreicht in der Regel eine viel größere Dicke als die oberirdischen Triebe, egal wo sie wachsen – im Wasser oder in einer anderen Umgebung. Im Zusammenhang mit dem Hauptzweck des Rhizoms – als Speicher für Reservestoffe zu dienen – steht die Parenchymatisierung seines Gewebes im Vordergrund Besonderheit Strukturen. Das Speichergewebe in typischen Rhizomen ist der Kern, der aus parenchymalen, runden, dünnwandigen Zellen mit kleinen Interzellularräumen dazwischen besteht. Der Kern in den Rhizomen überwiegt in seiner Massivität gegenüber anderen Geweben. Mechanische und leitfähige Gewebe, die dem Kern und der Rinde nachgeben, sind im Rhizom weniger ausgeprägt als in oberirdischen Trieben.
Die Struktur des Rhizoms als mehrjähriger Spross entsteht unter dem Einfluss verschiedener prägender Umweltfaktoren. Der Einfluss dieser Faktoren kann sowohl direkt als auch durch eine Korrelation in der Entwicklung pflanzlicher Organe wirken: So beeinflussen beispielsweise der Entwicklungsgrad der Blätter, die Bildung von Blütentrieben und die Ablagerung von Reservestoffen die Struktur des Rhizoms. Besonders auffällig ist die Abhängigkeit der Struktur des Rhizoms vom Entwicklungsgrad der Blätter auf ihnen.
Während des unterirdischen Wachstums, wenn am Rhizom nur schuppenartige kleine Blätter vorhanden sind, bildet sich im Xylemteil ein sehr schwaches Leitungssystem, das aus seltenen Gefäßen mit schmaler Höhle und primitiver Struktur besteht. Nachdem sich der oberirdische fruchttragende Spross mit normal großen Blättern zu entwickeln beginnt, nimmt die Aktivität des Kambiums zu und infolgedessen entwickelt sich Holz sowie die Anzahl und das Lumen der Gefäße, die eine perfektere Struktur aufweisen als diejenigen, die vor Beginn des Wachstums des oberirdischen Sprosses gebildet wurden.
Ein besonderes morphologisches Merkmal des Rhizoms sind sehr kurze Internodien. Offenbar steht die Bedeutungslosigkeit der Internodienlänge in engem Zusammenhang mit der schwachen Differenzierung der Blätter am Rhizom.
In mancher Hinsicht ähnelt eine andere Modifikation des unterirdischen Triebs, eine Knolle, in ihrer Morphologie dem Rhizom. Der Hauptunterschied zwischen einem Rhizom und einer Knolle besteht in der Existenzdauer: Das Rhizom ist ein mehrjähriger Trieb, während die Knolle normalerweise ein bis zwei Jahre existiert. Daher zeigten sich in der Knolle nur die Strukturmerkmale, die die Rhizome im ersten Stadium der Gewebebildung aufweisen. Das Vorherrschen des Parenchyms in der Knolle ist noch deutlicher als im Rhizom.
Der Parenchymatisierungsgrad des Knollengewebes ähnelt dem des Stängels von Sukkulenten. Ebenso wie bei den Stängeln von Sukkulenten ist die gesamte Masse des Knollengewebes mit Ausnahme der Haut (Periderm) ein Speichergewebe, das von unterentwickelten und primitiv aufgebauten Bündeln des Leitungssystems durchdrungen ist. Zusammen mit der Reduzierung des leitfähigen Gewebes in einer normalen Knolle wird in der Regel überhaupt kein mechanisches Gewebe gebildet. Die Gewebe, aus denen die Knolle besteht, sind einheitlich aufgebaut und nur für eine einzige Funktion geeignet: die Ablagerung von Reserven.

Ein typisches Beispiel für eine Knollenstruktur wäre eine Kartoffelknolle. Die Abbildung zeigt einen Fragment eines Querschnitts einer Knolle einer der Kartoffelsorten („Woltman“) aus ihrem Mittelteil. Zur einfacheren Darstellung der Gewebestruktur ist Stärke, die alle Zellen im Überfluss ausfüllt, nicht dargestellt. Das Knollenparenchym wird nur an einer Stelle von einem schmalen Streifen kleinerer Zellen, bestehend aus vier Gruppen, durchtrennt. Jede Gruppe enthält 1-3 Gefäße mit enger Höhle, der Rest des Gewebes besteht aus meristemartigen Zellen, zwischen denen sich mehrere Siebröhren befinden. Gefäße, die sich durch eine unterentwickelte Struktur auszeichnen, sind primitiv und einige von ihnen befinden sich in einem Zustand der Auslöschung und Zerstörung.
Dieses aus primitiven und unterentwickelten Gefäßelementen bestehende Gewebe befindet sich ringförmig in der gesamten Knolle und stellt einen zentralen Zylinder dar. Der primäre Kortex liegt außen, der Kern innen. In Knollen, die nur schuppenähnliche Blätter haben, wird durch die Aktivität des Kambiums kein Sekundärgewebe gebildet.
Wenn Sie einen Fehler finden, markieren Sie bitte einen Textabschnitt und klicken Sie Strg+Eingabetaste.
Stengel. Morphologie und Funktionen des Stammes. Stängel - länglicher Trieb große Pflanzen, die als mechanische Achse dient, übernimmt auch die Funktion einer leitenden und stützenden Basis für Blätter, Knospen und Blüten. Die Stängel erfüllen zwei Hauptfunktionen – Tragen und Führen. Stängel verbinden alle Teile des Triebs zu einem einzigen System. Dank des Stängels wird eine Vergrößerung der Pflanzenoberfläche durch deren Verzweigung, Bildung und eine günstigere Anordnung der Blätter und gewährleistet Zeugungsorgane. Der Stiel sorgt für die auf- und absteigende Bewegung von Wasser und darin gelösten Substanzen. Junge Triebe übernehmen die Funktion der Photosynthese. Modifizierte Stängel erfüllen auch zusätzliche Funktionen, beispielsweise Akkumulation und Speicherung Nährstoffe in mehrjährigen Stängeln, sowie Wasserspeicherung in spezialisierten Stängeln, Haken anderer Pflanzen, Schutz, Versorgung vegetative Vermehrung Pflanzen.
Pflanzenstämme sind sehr vielfältig und unterscheiden sich in vielerlei Hinsicht. Je nach Konsistenz sind die Stängel: krautig oder holzig, hohl (Stroh) oder gefüllt (mit Kern). Je nach Form des Querschnitts sind die Stängel rund oder zylindrisch (bei den meisten Pflanzenarten), tetraedrisch (Arten der Familien Glukhokropiv und Norichnikov), dreiflächig (Arten der Seggenfamilie), vielfältig (Arten von Quinoa, Sauerampfer), abgeflacht (z. B. komprimiertes Bluegrass, Binsenmitglied rein). Je nach der Art des Wachstums und der Platzierung im Raum werden folgende Arten von Stängeln unterschieden: aufrecht (starke, meist orthotrope Stängel mit gut definierten Internodien, die meisten Arten höherer Pflanzen) kriechend (meist plagiotrope Stängel wurzeln an den Knoten, mit gut definierten Internodien; zum Beispiel kriechend zäh, Erdbeeren, kriechender Hahnenfuß, Gänsefingerkraut) lockig (im Uhrzeigersinn um die Stütze drehen oder umgekehrt; zum Beispiel Hopfen, Ackerwinde). ed, Dodder, Bohnen) kletternd oder hartnäckig (Stängel mit schlecht entwickelten). mechanisches Gewebe mit Antennen verbunden; zum Beispiel Gurken, gewöhnlicher Kürbis, gewöhnliche Erbsen) kriechend (Stängel mit eher schwach entwickeltem mechanischem Gewebe, entlang der Bodenoberfläche kriechen: zum Beispiel nackte Hernie, liegende Bryozoen, kriechende St. sehr enge Knoten, kurze Internodien und Blätter, die in einer basalen Rosette gesammelt sind, bilden einen Blütenpfeil; zum Beispiel Wegerich, Löwenzahn, Taglilien, Schneeglöckchen). Vorbaumodifikationen.
Der modifizierte Stamm ist ein Organ, das dazu dient, Reservenährstoffe anzusammeln, die die Pflanze zum Überleben der Ruhephase benötigt. Darüber hinaus sind veränderte Stängel häufig an der Vermehrung und Verbreitung von Pflanzen beteiligt. Um dieser Gruppe zugeordnet zu werden, muss ein modifizierter Stängel alle Merkmale eines gewöhnlichen Stängels aufweisen, d. Der Unterschied zwischen dem modifizierten Stamm und dem normalen besteht darin, dass er nicht unbedingt über dem Boden wächst, sein Wachstum oft mehr oder weniger horizontal verläuft und er normalerweise ein Reservoir für Reservenährstoffe darstellt.
Es gibt sechs Haupttypen modifizierter Stängel: Knolle, Rhizom, Knolle, Zwiebel, Schnurrbart, Nachkommen
Die Entstehung von Primärgewebe und der primären anatomischen Struktur. Die anatomische Struktur des Pflanzenstamms wird durch seine Hauptfunktionen bestimmt. Es ist durch die Entwicklung mechanischer und leitfähiger Gewebe von Pflanzen gekennzeichnet. Darüber hinaus zeichnet sich der Stamm durch ein komplexes System von Meristemen aus – apikal, lateral und interkalar –, die sein langfristiges Wachstum und die Entstehung neuer Organe bestimmen.
Der Stamm einer Pflanze entsteht aus Apikalmeristems, aus denen drei Gewebeschichten unterschieden werden: Hautgewebe, Leitgewebe, Hauptgewebe. Diese Gewebe werden zunächst durch das sogenannte Primärmeristem (Promeristem) dargestellt, bestehend aus Protoderm, Prokambium und dem Hauptmeristem. Aus Prokambium entwickeln sich primär leitfähige Gewebe. Die ersten Elemente des Phloems differenzieren sich von den äußeren Zellen des Prokambiums, die sich zur Peripherie hin befinden. Das primäre Phloem besteht aus dünnwandigen, kurzlebigen länglichen Zellen und wird Protophloem genannt, seine äußeren Zellen können durch mechanische Fasern dargestellt werden. Die primären Elemente des Xylems – Tracheiden, seltener Gefäße mit ringförmiger und spiralförmiger Wandverdickung – entstehen später aus den inneren Zellen des Prokambiums und werden allgemein als Protoxylem definiert. Zu seiner Zusammensetzung gehören neben leitenden Elementen auch Parenchymzellen. Später differenziert sich nach innen eine für das Phloem mehr oder weniger typische Struktur vom Protophloem. Außerhalb des Protoxylems bildet sich ein Metaxylem, bestehend aus Tracheiden oder Tracheen mit dickeren verholzten Wänden.
Durch die Aktivität des Prokambiums und des restlichen Apexmeristems entsteht somit die Primärstruktur des Pflanzenstamms. Bei einkeimblättrigen Pflanzen differenziert sich das gesamte Prokambium in Elemente primärer Leitgewebe. Die Stängel einkeimblättriger, insbesondere krautiger (Getreide) Pflanzen haben im Vergleich zu den Stängeln zweikeimblättriger Pflanzen eine einfachere Struktur, sie zeichnen sich hauptsächlich durch eine Primärstruktur aus. Bei zweikeimblättrigen Pflanzen kommt es im mittleren Teil des Prokambialstrangs zur Kambiumbildung und die Bildung sekundärer leitfähiger Gewebe (Metaphloeme und Metaxyleme) beginnt, deren Volumen durch die Teilung der Kambialzellen zunimmt. Bündel mit Kambium ium, charakteristisch für Dikotyledonen, sind offen. Krautige Monokotyledonen zeichnen sich durch eine diffuse Verteilung der Leitbündel aus. Leitende Bündel sind geschlossen, kollateral und selten konzentrisch. Von den mechanischen Geweben ist das Sklerenchym am weitesten entwickelt; das Kollenchym kommt in einigen Pflanzen vor. Bei krautigen Monokotyledonen gibt es keine sekundäre Verdickung.